Identification and Characterization of “Candidatus Rickettsia Thierseensis”, a Novel Spotted Fever Group Rickettsia Species Detected in Austria

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tick Identification and DNA Extraction
2.2. PCRs of Rickettsial Genes
2.3. PCR Purification and Sequencing
2.4. Data Availability
3. Results
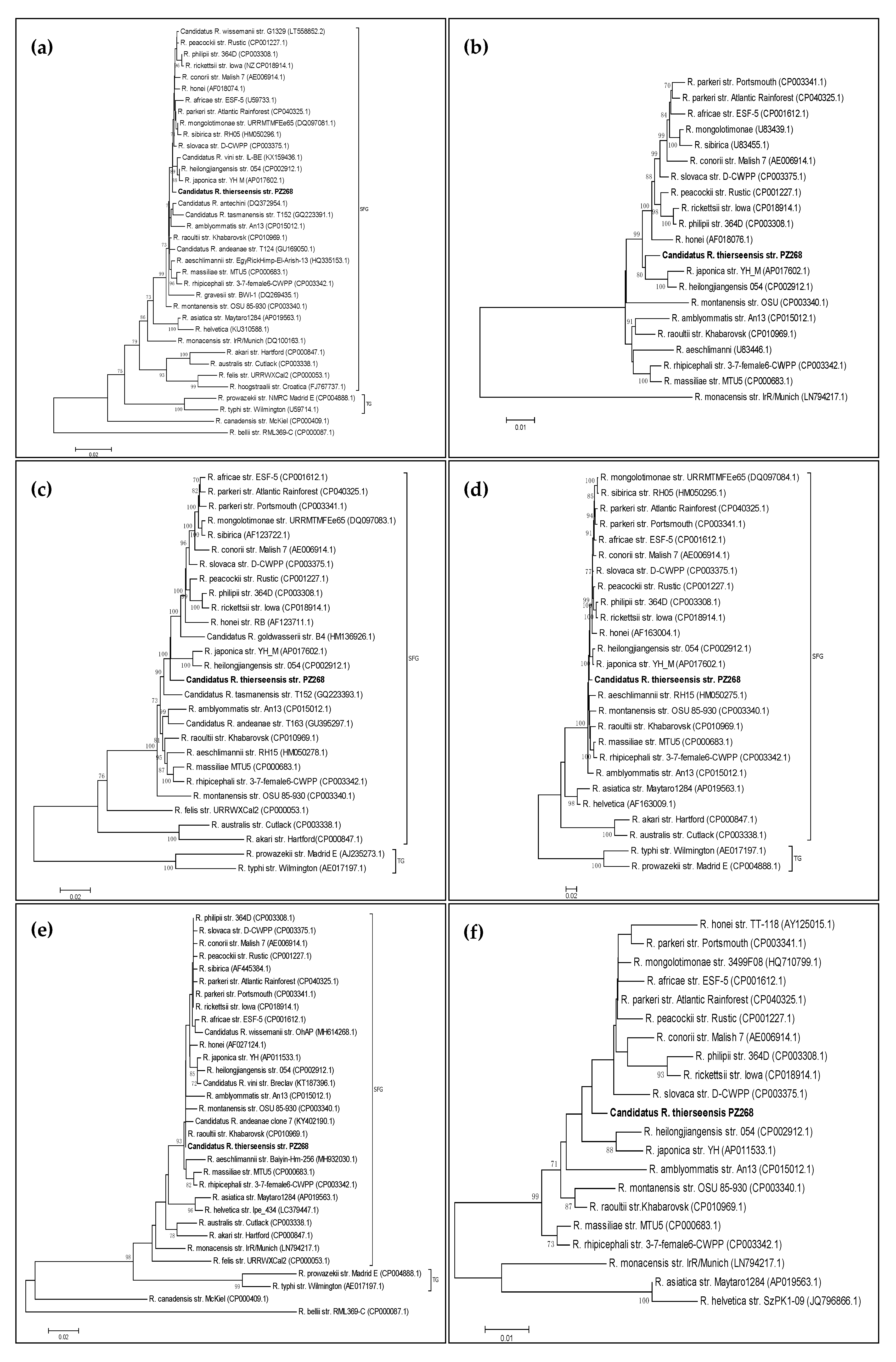
3.1. Phylogenetic Analysis of the gltA Gene
3.2. Phylogenetic Analysis of the ompA Gene
3.3. Phylogenetic Analysis of the ompB Gene
3.4. Phylogenetic Analysis of the sca4 Gene (Gene D)
3.5. Phylogenetic Analysis of the htrA Gene (17-kDa Antigen)
3.6. Phylogenetic Analysis of the 16S rRNA Gene
3.7. Phylogenetic Analysis of the 23S-5S Intergenic Spacer (IGS)
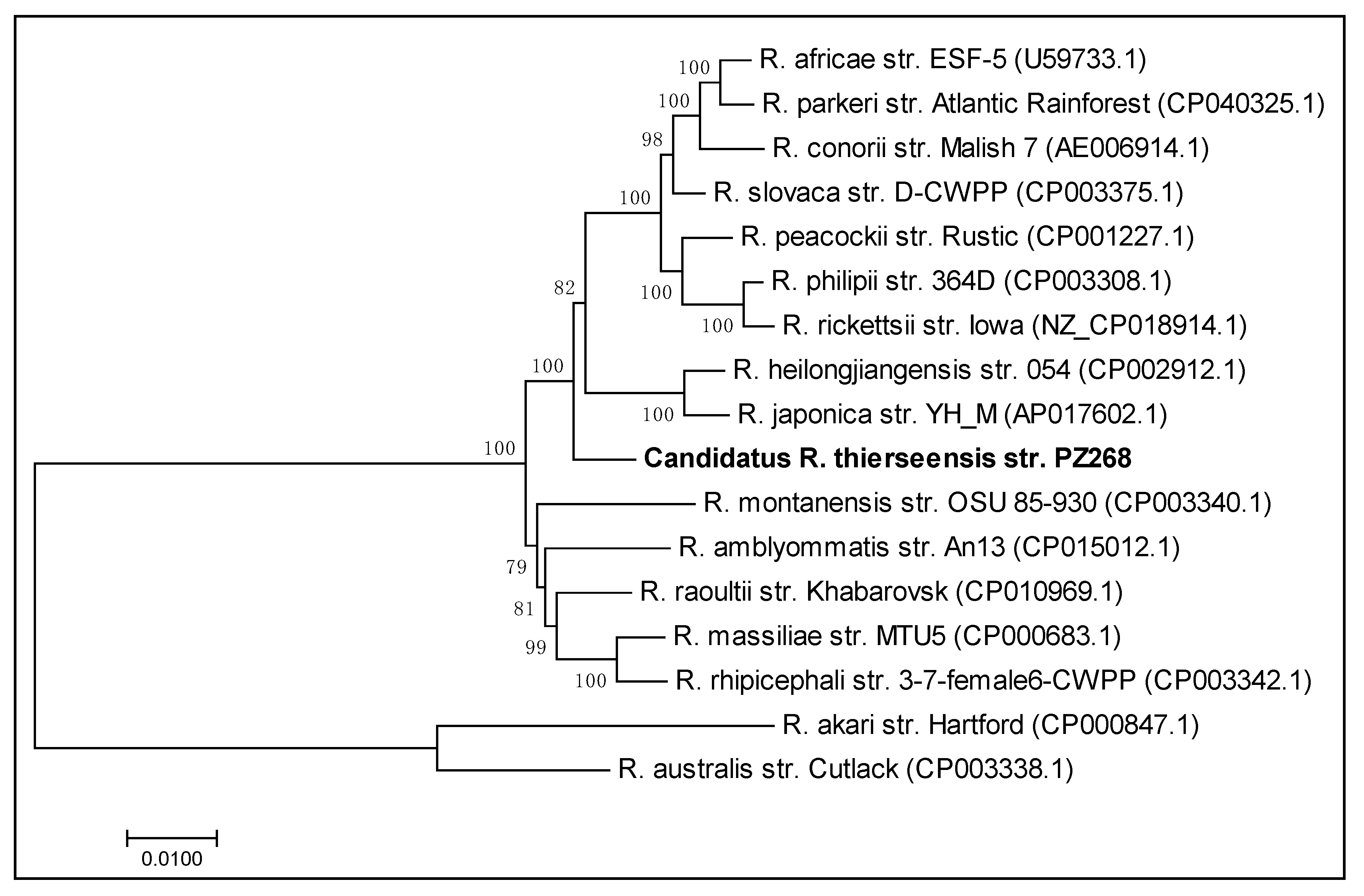
3.8. Multigenic Analysis of the Concatenated Sequences
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Diop, A.; El Karkouri, K.; Raoult, D.; Fournier, P.-E. Genome sequence-based criteria for demarcation and definition of species in the genus Rickettsia. Int. J. Syst. Evol. Microbiol. 2020, 70, 1738–1750. [Google Scholar] [CrossRef] [PubMed]
- Shpynov, S.N.; Fournier, P.E.; Pozdnichenko, N.N.; Gumenuk, A.S.; Skiba, A.A. New approaches in the systematics of rickettsiae. New Microbes New Infect. 2018, 23, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Paddock, C.D.; Raoult, D. Tick-borne rickettsioses around the world: Emerging diseases challenging old concepts. Clin. Microbiol. Rev. 2005, 18, 719–756. [Google Scholar] [CrossRef] [Green Version]
- Schötta, A.M.; Wijnveld, M.; Stockinger, H.; Stanek, G. Approaches for reverse line blot-based detection of microbial pathogens in Ixodes ricinus ticks collected in Austria and impact of the chosen method. Appl. Environ. Microbiol. 2017, 83, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, P.E.; Dumler, J.S.; Greub, G.; Zhang, J.; Wu, Y.; Raoult, D. Gene sequence-based criteria for identification of new Rickettsia isolates and description of Rickettsia heilongjiangensis sp. nov. J Clin Microbiol 2003, 41, 5456–5465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, W.C., 4th; Piesman, J. Phylogeny of hard- and soft-tick taxa (Acari: Ixodida) based on mitochondrial 16S rDNA sequences. Proc. Natl. Acad. Sci. USA 1994, 91, 10034–10038. [Google Scholar] [CrossRef] [Green Version]
- Labruna, M.B.; McBride, J.W.; Bouyer, D.H.; Camargo, L.M.A.; Camargo, E.P.; Walker, D.H. Molecular evidence for a spotted fever group Rickettsia species in the tick Amblyomma longirostre in Brazil. J. Med. Entomol. 2004, 41, 533–537. [Google Scholar] [CrossRef]
- Fournier, P.E.; Roux, V.; Raoult, D. Phylogenetic analysis of spotted fever group rickettsiae by study of the outer surface protein rOmpA. Int. J. Syst. Bacteriol. 1998, 48 Pt 3, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Sekeyova, Z.; Roux, V.; Raoult, D. Phylogeny of Rickettsia spp. inferred by comparing sequences of “gene D”, which encodes an intracytoplasmic protein. Int. J. Syst. Evol. Microbiol. 2001, 51, 1353–1360. [Google Scholar] [CrossRef] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletie, D.A.; Lane, D.J.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Jado, I.; Escudero, R.; Gil, H.; Jimenez-Alonso, M.I.; Sousa, R.; Garcia-Perez, A.L.; Rodriguez-Vargas, M.; Lobo, B.; Anda, P. Molecular method for identification of Rickettsia species in clinical and environmental samples. J. Clin. Microbiol. 2006, 44, 4572–4576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Welinder-Olsson, C.; Kjellin, E.; Vaht, K.; Jacobsson, S.; Wennerås, C. First case of human “Candidatus neoehrlichia mikurensis” infection in a febrile patient with chronic lymphocytic leukemia. J. Clin. Microbiol. 2010, 48, 1956–1959. [Google Scholar] [CrossRef] [Green Version]
- Schouls, L.M.; Van de Pol, I.; Rijpkema, S.G.T.; Schot, C.S. Detection and Identification of Ehrlichia, Borrelia burgdorferi Sensu Lato, and Bartonella Species in Dutch Ixodes ricinus Ticks. J. Clin. Microbiol. 1999, 37, 2215–2222. [Google Scholar] [CrossRef] [Green Version]
- Fukunaga, M.; Takahashi, Y.; Tsuruta, Y.; Matsushita, O.; Ralph, D.; McClelland, M.; Nakao, M. Genetic and phenotypic analysis of Borrelia miyamotoi sp. nov., isolated from the ixodid tick Ixodes persulcatus, the vector for Lyme disease in Japan. Int. J. Syst. Bacteriol. 1995, 45, 804–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gugliotta, J.L.; Goethert, H.K.; Berardi, V.P.; Telford, S.R., III. Meningoencephalitis from Borrelia miyamotoi in an Immunocompromised Patient. N. Engl. J. Med. 2013, 368, 240–245. [Google Scholar] [CrossRef] [Green Version]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, K.; Lindquist, O.; Påhlson, C. Association of Rickettsia helvetica with chronic perimyocarditis in sudden cardiac death. Lancet 1999, 354, 1169–1173. [Google Scholar] [CrossRef]
- Beati, L.; Peter, O.; Burgdorfer, W.; Aeschlimann, A.; Raoult, D. Confirmation that Rickettsia helvetica sp. nov. is a distinct species of the spotted fever group of rickettsiae. Int. J. Syst. Bacteriol. 1993, 43, 521–526. [Google Scholar] [CrossRef]
- Mediannikov, O.; Matsumoto, K.; Samoylenko, I.; Drancourt, M.; Roux, V.; Rydkina, E.; Davoust, B.; Tarasevich, I.; Brouqui, P.; Fournier, P.E. Rickettsia raoultii sp. nov., a spotted fever group rickettsia associated with Dermacentor ticks in Europe and Russia. Int. J. Syst. Evol. Microbiol. 2008, 58, 1635–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simser, J.A.; Palmer, A.T.; Fingerle, V.; Wilske, B.; Kurtti, T.J.; Munderloh, U.G. Rickettsia monacensis sp. nov., a spotted fever group rickettsia, from ticks (Ixodes ricinus) collected in a European city park. Appl. Environ. Microbiol. 2002, 68, 4559–4566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jado, I.; Oteo, J.A.; Aldámiz, M.; Gil, H.; Escudero, R.; Ibarra, V.; Portu, J.; Portillo, A.; Lezaun, M.J.; García-Amil, C.; et al. Rickettsia monacensis and human disease, Spain. Emerg. Infect. Dis. 2007, 13, 1405–1407. [Google Scholar] [CrossRef]
- Raoult, D.; Berbis, P.; Roux, V.; Xu, W.; Maurin, M. A new tick-transmitted disease due to Rickettsia slovaca. Lancet 1997, 350, 112–113. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene | Fragment | Primer Name | Primer Sequence (5′→3′) | Reference |
|---|---|---|---|---|
| gltA | I 1 | CS-78 | GCAAGTATCGGTGAGGATGTAAT | [7] |
| CS-323 | GCTTCCTTAAAATTCAATAAATCAGGAT | |||
| II 1 | CS-239 | GCTCTTCTCATCCTATGGCTATTAT | ||
| CS-1069 | CAGGGTCTTCGTGCATTTCTT | |||
| ompA | I 1 | 190-70 | ATGGCGAATATTTCTCCAAAA | [8] |
| 190-701 | GTTCCGTTAATGGCAGCATCT | |||
| II 1 | 190-3588 | AACAGTGAATGTAGGAGCAG | ||
| 190-4406 | ACTATACCCTCATCGTCATT | |||
| III | 190-4338 | TTCAGGAAACGACCGTACG | ||
| 190-5238 | ACTATTAAAGGCTAGGCTATT | |||
| IV | 190-5125 | GCGGTTACTTTAGCCAAAGG | ||
| 190-6013 | TCTTCTGCGTTGCATTACCG | |||
| V 1 | 190-5917 | TCAGGGAATAAAGGTCCTG | ||
| 190-6808 | CACGAACTTTCACACTACC | |||
| ompB | I 1 | M59 F | CCGCAGGGTTGGTAACTGC | [9] |
| 120-807 R | CCTTTTAGATTACCGCCTAA | |||
| II | 120-607 F | AATATCGGTGACGGTCAAGG | ||
| 120-1497 | CCTATATCGCCGGTAATT | |||
| III | 120-1378 | CCTATATCGCCGGTAATT | ||
| 120-2399 | CTTGTTTGTTTAATGTTACGGT | |||
| IV | 120-2113 | CGATGCTAACGTAGGTTCTT | ||
| 120-2988 | CCGGCTATACCGCCTGTAGT | |||
| V | 120-2788 | AAACAATAATCAAGGTACTGT | ||
| 120-3599 | TACTTCCGGTTACAGCAAAGT | |||
| VI | 120-3462 | CCACAGGAACTACAACCATT | ||
| 120-4346 | CGAAGAAGTAACGCTGACTT | |||
| VII 1 | 120-4232 | GGTTTCTCATTCTCTCTATATGG | ||
| 120-4879 | TTAGAAGTTTACACGGACTTTT | |||
| sca4 | I 1 | D1f | ATGAGTAAAGACGGTAACCT | [10] |
| D928r | AAGCTATTGCGTCATCTCCG | |||
| II | D767f | CGATGGTAGCATTAAAAGCT | ||
| D1390r | CTTGCTTTTCAGCAATATCAC | |||
| III | D1219f | CCAAATCTTCTTAATACAGC | ||
| D1876r | TAGTTTGTTCTGCCATAATC | |||
| IV | D1738f | GTATCTGAATTAAGCAATGCG | ||
| D2482r | CTATAACAGGATTAACAGCG | |||
| V 1 | D2338f | GATGCAGCGAGTGAGGCAGC | ||
| D3069r | TCAGCGTTGTGGAGGGGAAG | |||
| htrA | I 1 | 17k-5 | GCTTTACAAAATTCTAAAAACCATATA | [7] |
| 17k-3 | TGTCTATCAATTCACAACTTGCC | |||
| 16S rDNA | I | fD1 | AGAGTTTGATCCTGGCTCAG | [11] |
| rP1 | ACGGTTACCTTGTTACGACTT | |||
| 23S-5S IGS | I 1 | RCK/23-5-F | GATAGGTCRGRTGTGGAAGCAC | [12] |
| RCK/23-5-R | TCGGGAYGGGATCGTGTGTTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schötta, A.-M.; Wijnveld, M.; Höss, D.; Stanek, G.; Stockinger, H.; Markowicz, M. Identification and Characterization of “Candidatus Rickettsia Thierseensis”, a Novel Spotted Fever Group Rickettsia Species Detected in Austria. Microorganisms 2020, 8, 1670. https://doi.org/10.3390/microorganisms8111670
Schötta A-M, Wijnveld M, Höss D, Stanek G, Stockinger H, Markowicz M. Identification and Characterization of “Candidatus Rickettsia Thierseensis”, a Novel Spotted Fever Group Rickettsia Species Detected in Austria. Microorganisms. 2020; 8(11):1670. https://doi.org/10.3390/microorganisms8111670
Chicago/Turabian StyleSchötta, Anna-Margarita, Michiel Wijnveld, Dieter Höss, Gerold Stanek, Hannes Stockinger, and Mateusz Markowicz. 2020. "Identification and Characterization of “Candidatus Rickettsia Thierseensis”, a Novel Spotted Fever Group Rickettsia Species Detected in Austria" Microorganisms 8, no. 11: 1670. https://doi.org/10.3390/microorganisms8111670