Genetic and Neuroimaging Approaches to Understanding Post-Traumatic Stress Disorder

 ,
,  , ,
, ,
Abstract
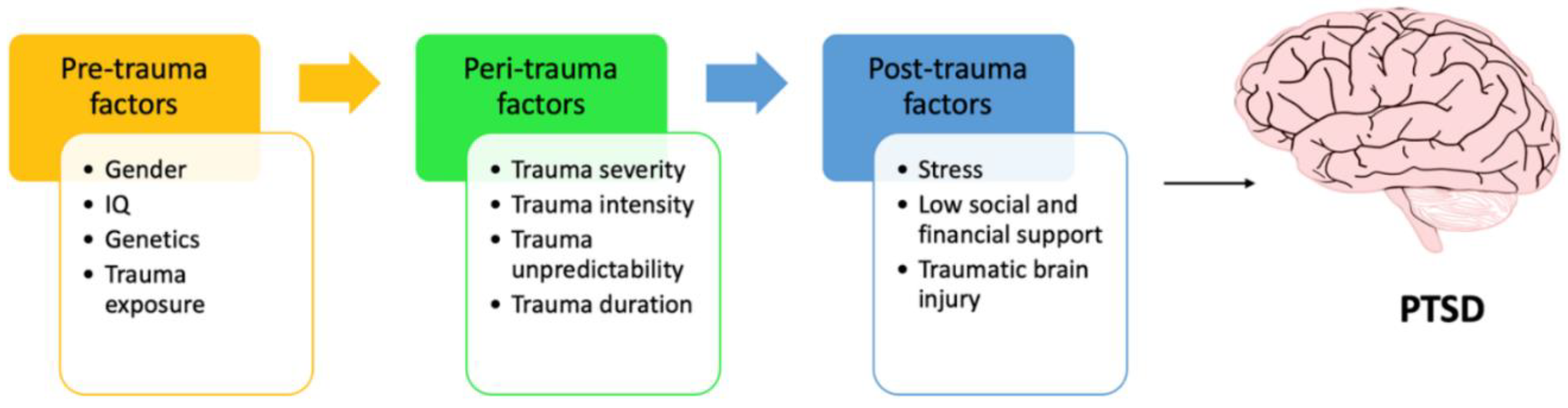
:1. Background
2. Neuroimaging in PTSD
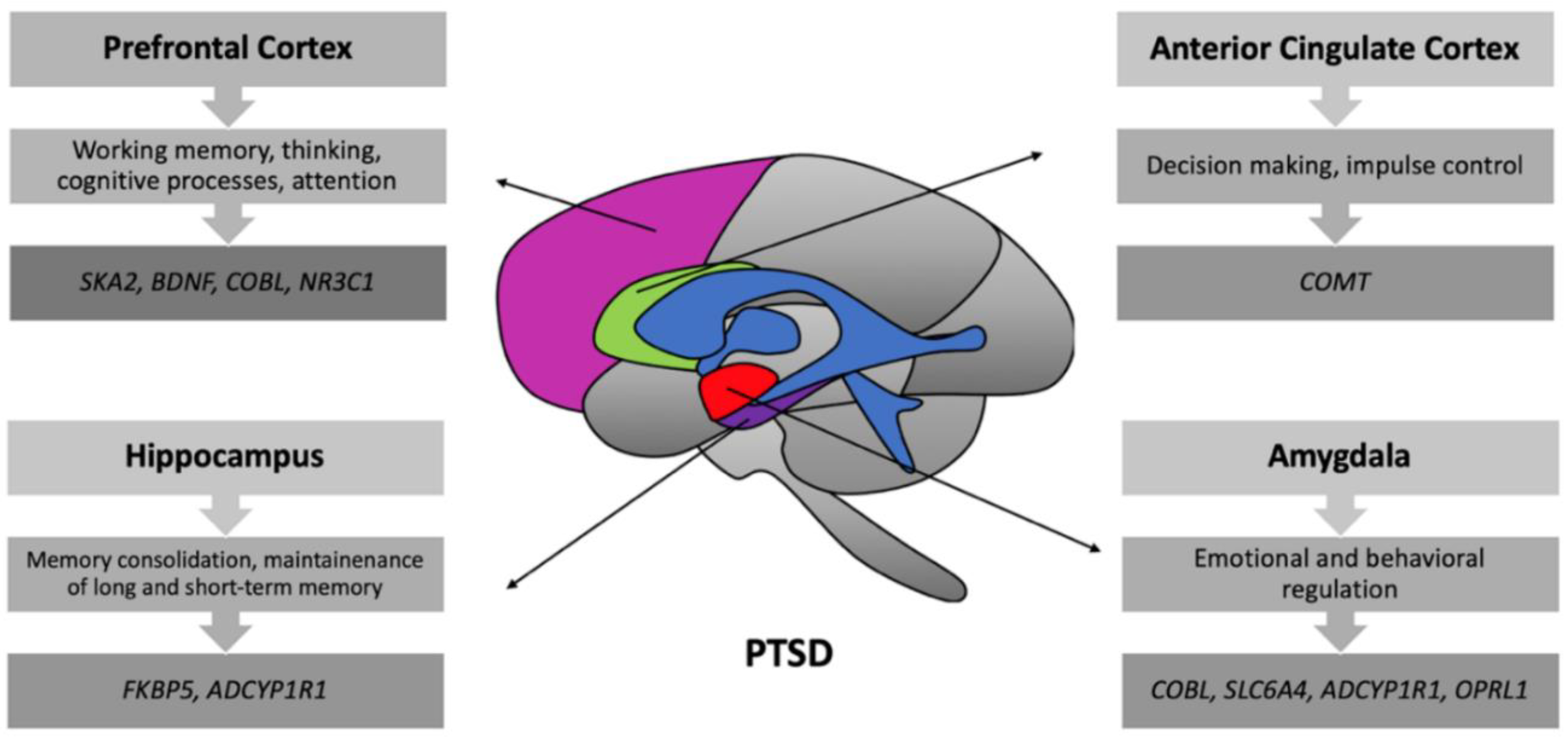
2.1. Structural Brain Changes
2.2. Functional Brain Changes
3. Genetic Differences in PTSD
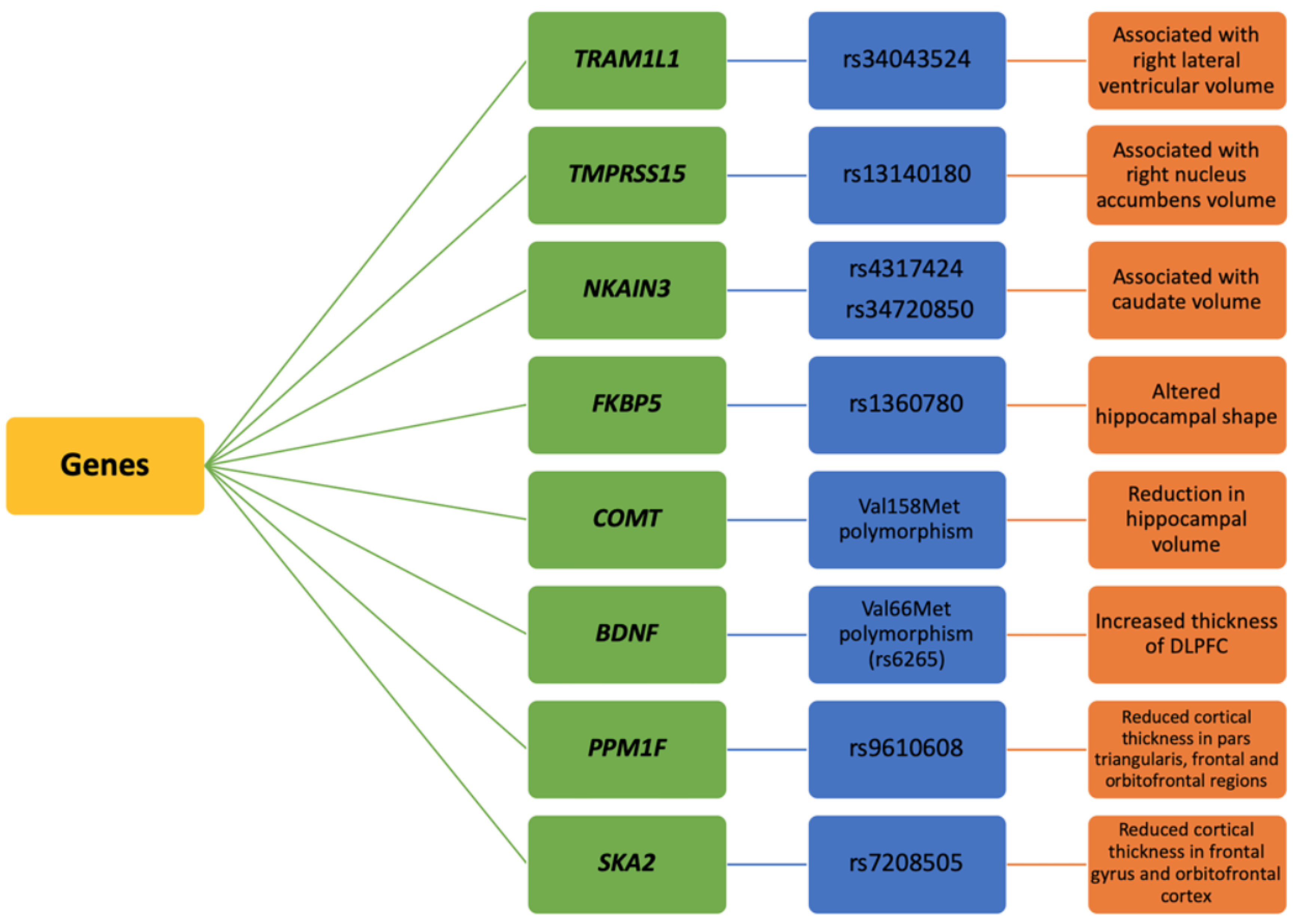
3.1. Genetics of Brain Morphology in PTSD
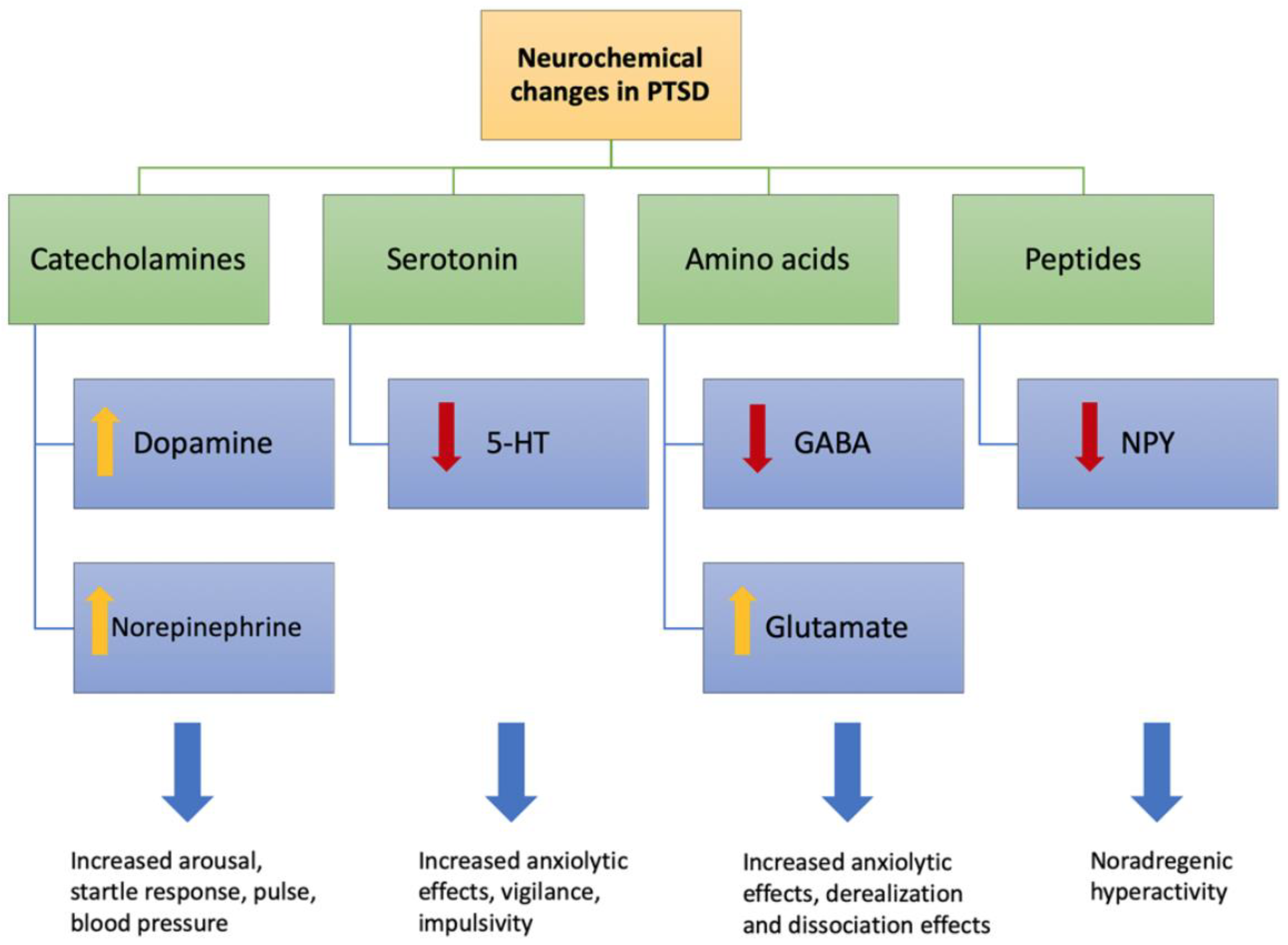
3.2. Genetics of Neuronal/Functional Changes in PTSD
4. Diagnostic Model Based on Imaging Genetics in PTSD
5. Potential Challenges and Future Perspective
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nardo, D.; Högberg, G.; Jonsson, C.; Jacobsson, H.; Hällström, T.; Pagani, M. Neurobiology of Sleep Disturbances in PTSD Patients and Traumatized Controls: MRI and SPECT Findings. Front. Psychiatry 2015, 6, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, S.; Iacoviello, B.M.; Charney, D.S. Risk Factors for the Development of Psychopathology Following Trauma. Curr. Psychiatry Rep. 2015, 17, 70. [Google Scholar] [CrossRef] [PubMed]
- Im, J.J.; Namgung, E.; Choi, Y.; Kim, J.Y.; Rhie, S.J.; Yoon, S. Molecular Neuroimaging in Posttraumatic Stress Disorder. Exp. Neurobiol. 2016, 25, 277–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fani, N.; Gutman, D.; Tone, E.B.; Almli, L.; Mercer, K.B.; Davis, J.; Glover, E.; Jovanovic, T.; Bradley, B.; Dinov, I.D.; et al. FKBP5 and attention bias for threat: Associations with hippocampal function and shape. JAMA Psychiatry 2013, 70, 392–400. [Google Scholar] [CrossRef] [Green Version]
- Fani, N.; King, T.Z.; Reiser, E.; Binder, E.B.; Jovanovic, T.; Bradley, B.; Ressler, K.J. FKBP5 genotype and structural integrity of the posterior cingulum. Neuropsychopharmacology 2014, 39, 1206–1213. [Google Scholar] [CrossRef] [Green Version]
- Schulz-Heik, R.J.; Schaer, M.; Eliez, S.; Hallmayer, J.F.; Lin, X.; Kaloupek, D.G.; Woodward, S.H. Catechol-O-Methyltransferase Val158Met Polymorphism Moderates Anterior Cingulate Volume in Posttraumatic Stress Disorder. Biol. Psychiatry 2011, 70, 1091–1096. [Google Scholar] [CrossRef]
- Lyoo, I.K.; Kim, J.E.; Yoon, S.J.; Hwang, J.; Bae, S.; Kim, D.J. The Neurobiological Role of the Dorsolateral Prefrontal Cortex in Recovery From Trauma: Longitudinal Brain Imaging Study Among Survivors of the South Korean Subway Disaster. Arch. Gen. Psychiatry 2011, 68, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Almli, L.M.; Srivastava, A.; Fani, N.; Kerley, K.; Mercer, K.B.; Feng, H.; Bradley, B.; Ressler, K.J. Follow-up and Extension of a Prior Genome-wide Association Study of Posttraumatic Stress Disorder: Gene x Environment Associations and Structural Magnetic Resonance Imaging in a Highly Traumatized African-American Civilian Population. Biol. Psychiatry 2014, 76, e3–e4. [Google Scholar] [CrossRef] [Green Version]
- Morey, R.A.; Hariri, A.R.; Gold, A.L.; Hauser, M.A.; Munger, H.J.; Dolcos, F.; McCarthy, G. Serotonin transporter gene polymorphisms and brain function during emotional distraction from cognitive processing in post-traumatic stress disorder. BMC Psychiatry 2011, 11, 76. [Google Scholar] [CrossRef] [Green Version]
- Stevens, J.S.; Almli, L.M.; Fani, N.; Gutman, D.A.; Bradley, B.; Norrholm, S.D.; Reiser, E.; Ely, T.D.; Dhanani, R.; Glover, E.M.; et al. PACAP receptor gene polymorphism impacts fear responses in the amygdala and hippocampus. Proc. Natl. Acad. Sci. USA 2014, 111, 3158–3163. [Google Scholar] [CrossRef] [Green Version]
- Andero, R.; Brothers, S.P.; Jovanovic, T.; Chen, Y.T.; Salah-Uddin, H.; Cameron, M.; Bannister, T.D.; Almli, L.; Stevens, J.S.; Bradley, B.; et al. Amygdala-dependent fear is regulated by Oprl1 in mice and humans with PTSD. Sci. Transl. Med. 2013, 5, ra73–ra188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeh, N.; Spielberg, J.M.; Logue, M.W.; Wolf, E.J.; Smith, A.K.; Lusk, J.; Hayes, J.P.; Sperbeck, E.; Milberg, W.P.; McGlinchey, R.E.; et al. SKA2 methylation is associated with decreased prefrontal cortical thickness and greater PTSD severity among trauma-exposed veterans. Mol. Psychiatry 2016, 21, 357–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schechter, D.S.; Moser, D.A.; Paoloni-Giacobino, A.; Stenz, L.; Gex-Fabry, M.; Aue, T.; Adouan, W.; Cordero, M.I.; Suardi, F.; Manini, A.; et al. Methylation of NR3C1 is related to maternal PTSD, parenting stress and maternal medial prefrontal cortical activity in response to child separation among mothers with histories of violence exposure. Front. Psychol. 2015, 6, 690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symms, M.; Jager, H.; Schmierer, K.; Yousry, T. A review of structural magnetic resonance neuroimaging. J. Neurol. Neurosurg. Psychiatry 2004, 75, 1235–1244. [Google Scholar] [CrossRef]
- Morey, R.A.; Gold, A.L.; LaBar, K.S.; Beall, S.K.; Brown, V.M.; Haswell, C.C.; Nasser, J.D.; Wagner, H.R.; McCarthy, G.; Mid-Atlantic, M.W. Amygdala volume changes in post-traumatic stress disorder in a large case-controlled veterans group. Arch. Gen. Psychiatry 2012, 69, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Neylan, T.C.; Mueller, S.G.; Lenoci, M.; Truran, D.; Marmar, C.R.; Weiner, M.W.; Schuff, N. Magnetic Resonance Imaging of Hippocampal Subfields in Posttraumatic Stress Disorder. Arch. Gen. Psychiatry 2010, 67, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Hughes, K.C.; Shin, L.M. Functional neuroimaging studies of post-traumatic stress disorder. Expert Rev. Neurother. 2011, 11, 275–285. [Google Scholar] [CrossRef]
- Herringa, R.; Phillips, M.; Almeida, J.; Insana, S.; Germain, A. Post-traumatic stress symptoms correlate with smaller subgenual cingulate, caudate, and insula volumes in unmedicated combat veterans. Psychiatry Res. 2012, 203, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Morey, R.A.; Haswell, C.C.; Hooper, S.R.; De Bellis, M.D. Amygdala, Hippocampus, and Ventral Medial Prefrontal Cortex Volumes Differ in Maltreated Youth with and without Chronic Posttraumatic Stress Disorder. Neuropsychopharmacology 2016, 41, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Kühn, S.; Gallinat, J. Gray Matter Correlates of Posttraumatic Stress Disorder: A Quantitative Meta-Analysis. Biol. Psychiatry 2013, 73, 70–74. [Google Scholar] [CrossRef]
- O’Doherty, D.C.M.; Tickell, A.; Ryder, W.; Chan, C.; Hermens, D.F.; Bennett, M.R.; Lagopoulos, J. Frontal and subcortical grey matter reductions in PTSD. Psychiatry Res. Neuroimaging 2017, 266, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vermetten, E.; Vythilingam, M.; Southwick, S.M.; Charney, D.S.; Bremner, J.D. Long-Term Treatment with Paroxetine Increases Verbal Declarative Memory and Hippocampal Volume in Posttraumatic Stress Disorder. Biol. Psychiatry 2003, 54, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Tan, Q.; Yin, H.; Zhang, X.; Huan, Y.; Tang, L.; Wang, H.; Xu, J.; Li, L. Decreased gray matter volume in the left hippocampus and bilateral calcarine cortex in coal mine flood disaster survivors with recent onset PTSD. Psychiatry Res. Neuroimaging 2011, 192, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, M.W.; Shenton, M.E.; Ciszewski, A.; Kasai, K.; Lasko, N.B.; Orr, S.P.; Pitman, R.K. Smaller hippocampal volume predicts pathologic vulnerability to psychological trauma. Nat. Neurosci. 2002, 5, 1242–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Li, Y.-J.; Luo, E.-P.; Lu, H.-B.; Yin, H. Cortical Thinning in Patients with Recent Onset Post-Traumatic Stress Disorder after a Single Prolonged Trauma Exposure. PLoS ONE 2012, 7, e39025. [Google Scholar] [CrossRef]
- Qi, S.; Mu, Y.; Liu, K.; Zhang, J.; Huan, Y.; Tan, Q.; Shi, M.; Wang, Q.; Chen, Y.; Wang, H.; et al. Cortical inhibition deficits in recent onset PTSD after a single prolonged trauma exposure. Neuroimage Clin. 2013, 3, 226–233. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Lei, D.; Li, L.; Huang, X.; Suo, X.; Xiao, F.; Kuang, W.; Li, J.; Bi, F.; Lui, S.; et al. White Matter Abnormalities in Post-traumatic Stress Disorder Following a Specific Traumatic Event. EBioMedicine 2016, 4, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Schuff, N.; Zhang, Y.; Zhan, W.; Lenoci, M.; Ching, C.; Boreta, L.; Mueller, S.G.; Wang, Z.; Marmar, C.R.; Weiner, M.W.; et al. Patterns Of Altered Cortical Perfusion And Diminished Subcortical Integrity In Posttraumatic Stress Disorder: A MRI Study. NeuroImage 2011, 54 (Suppl. 1), S62–S68. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, B.; Feng, N.; Pu, H.; Zhang, X.; Lu, H.; Yin, H. Perfusion Deficits and Functional Connectivity Alterations in Memory-Related Regions of Patients with Post-Traumatic Stress Disorder. PLoS ONE 2016, 11, e0156016. [Google Scholar] [CrossRef]
- Ding, A.Y.; Li, Q.; Zhou, I.Y.; Ma, S.J.; Tong, G.; McAlonan, G.M.; Wu, E.X. MR Diffusion Tensor Imaging Detects Rapid Microstructural Changes in Amygdala and Hippocampus Following Fear Conditioning in Mice. PLoS ONE 2013, 8, e51704. [Google Scholar] [CrossRef] [Green Version]
- Lei, D.; Li, L.; Li, L.; Suo, X.; Huang, X.; Lui, S.; Li, J.; Bi, F.; Kemp, G.J.; Gong, Q. Microstructural Abnormalities in Children with Post-traumatic Stress Disorder: A Diffusion Tensor Imaging Study at 3.0T. Sci. Rep. 2015, 5, 8933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, O.; Yamasue, H.; Kasai, K.; Yamada, H.; Aoki, S.; Iwanami, A.; Ohtani, T.; Masutani, Y.; Kato, N.; Ohtomo, K. Voxel-based diffusion tensor analysis reveals aberrant anterior cingulum integrity in post-traumatic stress disorder due to terrorism. Psychiatry Res. Neuroimaging 2006, 146, 231–242. [Google Scholar] [CrossRef]
- Bing, X.; Ming-Guo, Q.; Ye, Z.; Jing-Na, Z.; Min, L.; Han, C.; Yu, Z.; Jia-jia, Z.; Jian, W.; Wei, C.; et al. Alterations in the cortical thickness and the amplitude of low-frequency fluctuation in patients with post-traumatic stress disorder. Brain Res. 2013, 1490, 225–232. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhang, R.; Li, K.; Qi, R.; Zhang, Z.; Huang, Q.; Lu, G. Altered cortical and subcortical local coherence in PTSD: Evidence from resting-state fMRI. Acta Radiol. 2015, 56, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Rabinak, C.A.; Angstadt, M.; Welsh, R.C.; Kenndy, A.E.; Lyubkin, M.; Martis, B.; Phan, K.L. Altered Amygdala Resting-State Functional Connectivity in Post-Traumatic Stress Disorder. Front. Psychiatry 2011, 2, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misaki, M.; Phillips, R.; Zotev, V.; Wong, C.-K.; Wurfel, B.E.; Krueger, F.; Feldner, M.; Bodurka, J. Connectome-wide investigation of altered resting-state functional connectivity in war veterans with and without post-traumatic stress disorder. Neuroimage Clin. 2018, 17, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, J.; Zhang, J.; Zhan, W.; Li, L.; Wu, M.; Huang, H.; Zhu, H.; Kemp, G.J.; Gong, Q. Altered resting-state functional activity in post-traumatic stress disorder: A quantitative meta-analysis. Sci. Rep. 2016, 6, 27131. [Google Scholar] [CrossRef] [Green Version]
- White, S.F.; Costanzo, M.E.; Blair, J.R.; Roy, M.J. PTSD symptom severity is associated with increased recruitment of top-down attentional control in a trauma-exposed sample. Neuroimage Clin. 2015, 7, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Blair, K.S.; Vythilingam, M.; Crowe, S.L.; McCaffrey, D.E.; Ng, P.; Wu, C.C.; Scaramozza, M.; Mondillo, K.; Pine, D.S.; Charney, D.S.; et al. Cognitive control of attention is differentially affected in trauma-exposed individuals with and without post-traumatic stress disorder. Psychol. Med. 2013, 43, 85–95. [Google Scholar] [CrossRef]
- Hayes, J.P.; LaBar, K.S.; Petty, C.M.; McCarthy, G.; Morey, R.A. Alterations in the neural circuitry for emotion and attention associated with post-traumatic stress symptomatology. Psychiatry Res. 2009, 172, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Bryant, R.A.; Felmingham, K.L.; Kemp, A.H.; Barton, M.; Peduto, A.S.; Rennie, C.; Gordon, E.; Williams, L.M. Neural Networks of Information Processing in Posttraumatic Stress Disorder: A Functional Magnetic Resonance Imaging Study. Biol. Psychiatry 2005, 58, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Landré, L.; Destrieux, C.; Andersson, F.; Barantin, L.; Quidé, Y.; Tapia, G.; Jaafari, N.; Clarys, D.; Gaillard, P.; Isingrini, M.; et al. Working memory processing of traumatic material in women with post-traumatic stress disorder. J. Psychiatry Neurosci. Jpn 2012, 37, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nejati, V.; Salehinejad, M.A.; Sabayee, A. Impaired working memory updating affects memory for emotional and non-emotional materials the same way: Evidence from post-traumatic stress disorder (PTSD). Cogn. Process. 2018, 19, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Pauly, K.; Kellermann, T.; Seiferth, N.Y.; Reske, M.; Backes, V.; Stöcker, T.; Shah, N.J.; Amunts, K.; Kircher, T.; et al. Gender differences in the cognitive control of emotion: An fMRI study. Neuropsychologia 2007, 45, 2744–2754. [Google Scholar] [CrossRef]
- Gamer, M.; Büchel, C. Amygdala Activation Predicts Gaze toward Fearful Eyes. J. Neurosci. 2009, 29, 9123. [Google Scholar] [CrossRef] [Green Version]
- Drobyshevsky, A.; Baumann, S.B.; Schneider, W. A Rapid fMRI Task Battery for Mapping of Visual, Motor, Cognitive and Emotional Function. NeuroImage 2006, 31, 732–744. [Google Scholar] [CrossRef] [Green Version]
- van Rooij, S.J.H.; Kennis, M.; Vink, M.; Geuze, E. Predicting Treatment Outcome in PTSD: A Longitudinal Functional MRI Study on Trauma-Unrelated Emotional Processing. Neuropsychopharmacology 2016, 41, 1156–1165. [Google Scholar] [CrossRef]
- Falconer, E.; Bryant, R.; Felmingham, K.L.; Kemp, A.H.; Gordon, E.; Peduto, A.; Olivieri, G.; Williams, L.M. The neural networks of inhibitory control in post-traumatic stress disorder. J. Psychiatry Neurosci. Jpn. 2008, 33, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Jovanovic, T.; Ressler, K.J. How the Neurocircuitry and Genetics of Fear Inhibition May Inform Our Understanding of PTSD. Am. J. Psychiatry 2010, 167, 648–662. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, S.; Niemeyer, H.; Engel, S.; Cwik, J.C.; Laufer, S.; Klusmann, H.; Knaevelsrud, C. HPA axis regulation in post-traumatic stress disorder: A meta-analysis focusing on potential moderators. Neurosci. Biobehav. Rev. 2019, 100, 35–57. [Google Scholar] [CrossRef]
- Hauer, D.; Kaufmann, I.; Strewe, C.; Briegel, I.; Campolongo, P.; Schelling, G. The role of glucocorticoids, catecholamines and endocannabinoids in the development of traumatic memories and post-traumatic stress symptoms in survivors of critical illness. Neurobiol. Learn. Mem. 2014, 112, 68–74. [Google Scholar] [CrossRef]
- Binder, E.B.; Bradley, R.G.; Liu, W.; Epstein, M.P.; Deveau, T.C.; Mercer, K.B.; Tang, Y.; Gillespie, C.F.; Heim, C.M.; Nemeroff, C.B.; et al. Association of FKBP5 Polymorphisms and Childhood Abuse With Risk of Posttraumatic Stress Disorder Symptoms in Adults. J. Am. Med. Assoc. 2008, 299, 1291–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenen, K.C.; Uddin, M. FKBP5 Polymorphisms Modify the Effects of Childhood Trauma. Neuropsychopharmacology 2010, 35, 1623–1624. [Google Scholar] [CrossRef] [PubMed]
- Sah, R.; Geracioti, T.D. Neuropeptide Y and post-traumatic stress disorder. Mol. Psychiatry 2013, 18, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Pivac, N.; Knezevic, J.; Kozaric-Kovacic, D.; Dezeljin, M.; Mustapic, M.; Rak, D.; Matijevic, T.; Pavelic, J.; Muck-Seler, D. Monoamine oxidase (MAO) intron 13 polymorphism and platelet MAO-B activity in combat-related post-traumatic stress disorder. J. Affect. Disord. 2007, 103, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.C.; Agrawal, A.; Pergadia, M.L.; Lynskey, M.T.; Todorov, A.A.; Wang, J.C.; Todd, R.D.; Martin, N.G.; Heath, A.C.; Goate, A.M.; et al. Association of childhood trauma exposure and GABRA2 polymorphisms with risk of post-traumatic stress disorder in adults. Mol. Psychiatry 2009, 14, 234–235. [Google Scholar] [CrossRef] [PubMed]
- Amstadter, A.B.; Koenen, K.C.; Ruggiero, K.J.; Acierno, R.; Galea, S.; Kilpatrick, D.G.; Gelernter, J. Variant in RGS2 Moderates Posttraumatic Stress Symptoms following Potentially Traumatic Event Exposure. J. Anxiety Disord. 2009, 23, 369–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logue, M.W.; Baldwin, C.; Guffanti, G.; Melista, E.; Wolf, E.J.; Reardon, A.F.; Uddin, M.; Wildman, D.; Galea, S.; Koenen, K.C.; et al. A genome-wide association study of post-traumatic stress disorder identifies the retinoid-related orphan receptor alpha (RORA) gene as a significant risk locus. Mol. Psychiatry 2013, 18, 937–942. [Google Scholar] [CrossRef] [Green Version]
- Amstadter, A.B.; Sumner, J.A.; Acierno, R.; Ruggiero, K.J.; Koenen, K.C.; Kilpatrick, D.G.; Galea, S.; Gelernter, J. Support for association of RORA variant and post traumatic stress symptoms in a population-based study of hurricane exposed adults. Mol. Psychiatry 2013, 18, 1148–1149. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.W.; Wolf, E.J.; Logue, M.W.; Baldwin, C.T. The retinoid-related orphan receptor alpha (RORA) gene and fear-related psychopathology. J. Affect. Disord. 2013, 151, 702–708. [Google Scholar] [CrossRef] [Green Version]
- Nievergelt, C.M.; Maihofer, A.X.; Mustapic, M.; Yurgil, K.A.; Schork, N.J.; Miller, M.W.; Logue, M.W.; Geyer, M.A.; Risbrough, V.B.; O’Connor, D.T.; et al. Genomic predictors of combat stress vulnerability and resilience in U.S. Marines: A genome-wide association study across multiple ancestries implicates PRTFDC1 as a potential PTSD gene. Psychoneuroendocrinology 2015, 51, 459–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, M.B.; Chen, C.-Y.; Ursano, R.J.; Cai, T.; Gelernter, J.; Heeringa, S.G.; Jain, S.; Jensen, K.P.; Maihofer, A.X.; Mitchell, C.; et al. Genome-wide Association Studies of Posttraumatic Stress Disorder in 2 Cohorts of US Army Soldiers. JAMA Psychiatry 2016, 73, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.C.; Heath, A.C.; Lynskey, M.T.; Agrawal, A.; Henders, A.K.; Bowdler, L.M.; Todorov, A.A.; Madden, P.A.F.; Moore, E.; Degenhardt, L.; et al. PTSD risk associated with a functional DRD2 polymorphism in heroin-dependent cases and controls is limited to amphetamine-dependent individuals. Addict. Biol. 2014, 19, 700–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelernter, J.; Sun, N.; Polimanti, R.; Pietrzak, R.; Levey, D.F.; Bryois, J.; Lu, Q.; Hu, Y.; Li, B.; Radhakrishnan, K.; et al. Genome-wide association study of post-traumatic stress disorder reexperiencing symptoms in >165,000 US veterans. Nat. Neurosci. 2019, 22, 1394–1401. [Google Scholar] [CrossRef]
- Freeman, T.; Roca, V.; Guggenheim, F.; Kimbrell, T.; Griffin, W.S.T. Neuropsychiatric Associations of Apolipoprotein E Alleles in Subjects With Combat-Related Posttraumatic Stress Disorder. J. Neuropsychiatry Clin. Neurosci. 2005, 17, 541–543. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Zuloaga, D.G.; Bidiman, E.; Marzulla, T.; Weber, S.; Wahbeh, H.; Raber, J. ApoE2 Exaggerates PTSD-Related Behavioral, Cognitive, and Neuroendocrine Alterations. Neuropsychopharmacology 2015, 40, 2443–2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morey, R.A.; Davis, S.L.; Garrett, M.E.; Haswell, C.C.; Mid-Atlantic, M.W.; Marx, C.E.; Beckham, J.C.; McCarthy, G.; Hauser, M.A.; Ashley-Koch, A.E. Genome-wide association study of subcortical brain volume in PTSD cases and trauma-exposed controls. Transl. Psychiatry 2017, 7, 1265. [Google Scholar] [CrossRef] [PubMed]
- Gorokhova, S.; Bibert, S.; Geering, K.; Heintz, N. A novel family of transmembrane proteins interacting with β subunits of the Na,K-ATPase. Hum. Mol. Genet. 2007, 16, 2394–2410. [Google Scholar] [CrossRef] [Green Version]
- Fani, N.; King, T.Z.; Shin, J.; Srivastava, A.; Brewster, R.C.; Jovanovic, T.; Bradley, B.; Ressler, K.J. Structural and functional connectivity in post-traumatic stress disorder: Associations with FKBP5. Depress Anxiety 2016, 33, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Hartley, C.A.; Phelps, E.A. Changing fear: The neurocircuitry of emotion regulation. Neuropsychopharmacology 2010, 35, 136–146. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.P.; Logue, M.W.; Reagan, A.; Salat, D.; Wolf, E.J.; Sadeh, N.; Spielberg, J.M.; Sperbeck, E.; Hayes, S.M.; McGlinchey, R.E.; et al. COMT Val158Met polymorphism moderates the association between PTSD symptom severity and hippocampal volume. J. Psychiatry Neurosci. 2017, 42, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, D.R.; Morrison, F.G.; Wolf, E.J.; Logue, M.W.; Fortier, C.B.; Salat, D.H.; Fonda, J.R.; Stone, A.; Schichman, S.; Milberg, W.; et al. The PPM1F gene moderates the association between PTSD and cortical thickness. J. Affect. Disord. 2019, 259, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, X.-X.; Hu, X.-Z. Post-traumatic stress disorder risk and brain-derived neurotrophic factor Val66Met. World J. Psychiatry 2016, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.J.; Nilsson, E.K.; Jacobsson, J.A.; Stein, D.J.; Fredriksson, R.; Lind, L.; Schiöth, H.B. BDNF Polymorphisms Are Linked to Poorer Working Memory Performance, Reduced Cerebellar and Hippocampal Volumes and Differences in Prefrontal Cortex in a Swedish Elderly Population. PLoS ONE 2014, 9, e82707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumner, J.A.; Sheridan, M.A.; Drury, S.S.; Esteves, K.C.; Walsh, K.; Koenen, K.C.; McLaughlin, K.A. Variation in CACNA1C is Associated with Amygdala Structure and Function in Adolescents. J. Child Adolesc. Psychopharmacol. 2015, 25, 701–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzyzewska, I.M.; Ensink, J.B.M.; Nawijn, L.; Mul, A.N.; Koch, S.B.; Venema, A.; Shankar, V.; Frijling, J.L.; Veltman, D.J.; Lindauer, R.J.L.; et al. Genetic variant in CACNA1C is associated with PTSD in traumatized police officers. Eur. J. Hum. Genet. 2018, 26, 247–257. [Google Scholar] [CrossRef]
- Sherin, J.E.; Nemeroff, C.B. Post-traumatic stress disorder: The neurobiological impact of psychological trauma. Dialogues Clin. Neurosci. 2011, 13, 263–278. [Google Scholar]
- Berumen, L.C.; Rodríguez, A.; Miledi, R.; García-Alcocer, G. Serotonin receptors in hippocampus. Sci. World J. 2012, 2012, 823493. [Google Scholar] [CrossRef] [Green Version]
- Yohn, C.N.; Gergues, M.M.; Samuels, B.A. The role of 5-HT receptors in depression. Mol. Brain 2017, 10, 28. [Google Scholar] [CrossRef]
- Kimbrel, N.A.; Morissette, S.B.; Meyer, E.C.; Chrestman, R.; Jamroz, R.; Silvia, P.J.; Beckham, J.C.; Young, K.A. Effect of the 5-HTTLPR polymorphism on post-traumatic stress disorder, depression, anxiety, and quality of life among Iraq and Afghanistan veterans. Anxiety Stress Coping 2015, 28, 456–466. [Google Scholar] [CrossRef] [Green Version]
- Gressier, F.; Calati, R.; Balestri, M.; Marsano, A.; Alberti, S.; Antypa, N.; Serretti, A. The 5-HTTLPR Polymorphism and Posttraumatic Stress Disorder: A Meta-Analysis. J. Trauma. Stress 2013, 26, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Mellman, T.A.; Alim, T.; Brown, D.D.; Gorodetsky, E.; Buzas, B.; Lawson, W.B.; Goldman, D.; Charney, D.S. Serotonin polymorphisms and post-traumatic stress disorder in a trauma exposed African American population. Depress Anxiety 2009, 26, 993–997. [Google Scholar] [CrossRef]
- Drury, S.S.; Theall, K.P.; Keats, B.J.B.; Scheeringa, M. The Role of the Dopamine Transporter (DAT) in the Development of PTSD in Preschool Children. J. Trauma. Stress 2009, 22, 534–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.-C.; Koenen, K.C.; Galea, S.; Aiello, A.E.; Soliven, R.; Wildman, D.E.; Uddin, M. Molecular variation at the SLC6A3 locus predicts lifetime risk of PTSD in the Detroit Neighborhood Health Study. PLoS ONE 2012, 7, e39184. [Google Scholar] [CrossRef]
- Valente, N.L.M.; Vallada, H.; Cordeiro, Q.; Miguita, K.; Bressan, R.A.; Andreoli, S.B.; Mari, J.J.; Mello, M.F. Candidate-Gene Approach in Posttraumatic Stress Disorder After Urban Violence: Association Analysis of the Genes Encoding Serotonin Transporter, Dopamine Transporter, and BDNF. J. Mol. Neurosci. 2011, 44, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Lonsdorf, T.B.; Weike, A.I.; Nikamo, P.; Schalling, M.; Hamm, A.O.; Öhman, A. Genetic Gating of Human Fear Learning and Extinction: Possible Implications for Gene-Environment Interaction in Anxiety Disorder. Psychol. Sci. 2009, 20, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Holmes, S.E.; Girgenti, M.J.; Davis, M.T.; Pietrzak, R.H.; DellaGioia, N.; Nabulsi, N.; Matuskey, D.; Southwick, S.; Duman, R.S.; Carson, R.E.; et al. Altered metabotropic glutamate receptor 5 markers in PTSD: In vivo and postmortem evidence. Proc. Natl. Acad. Sci. USA 2017, 114, 8390–8395. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, G.; Liu, Y. The molecular physiology of CRH neurons. Front. Neuroendocr. 2012, 33, 67–84. [Google Scholar] [CrossRef] [Green Version]
- Dedic, N.; Kühne, C.; Gomes, K.S.; Hartmann, J.; Ressler, K.J.; Schmidt, M.V.; Deussing, J.M. Deletion of CRH From GABAergic Forebrain Neurons Promotes Stress Resilience and Dampens Stress-Induced Changes in Neuronal Activity. Front. Neurosci. 2019, 13, 986. [Google Scholar] [CrossRef]
- Reichmann, F.; Holzer, P. Neuropeptide Y: A stressful review. Neuropeptides 2016, 55, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Sweis, B.M.; Veverka, K.K.; Dhillon, E.S.; Urban, J.H.; Lucas, L.R. Individual differences in the effects of chronic stress on memory: Behavioral and neurochemical correlates of resiliency. Neuroscience 2013, 246, 142–159. [Google Scholar] [CrossRef] [PubMed]
- Rasmusson, A.M.; Hauger, R.L.; Morgan, C.A.; Bremner, J.D.; Charney, D.S.; Southwick, S.M. Low baseline and yohimbine-stimulated plasma neuropeptide Y (NPY) levels in combat-related PTSD. Biol. Psychiatry 2000, 47, 526–539. [Google Scholar] [CrossRef]
- Qiao, L.-Y. Neurotrophin signaling and visceral hypersensitivity. Front. Biol. 2014, 9, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Anna, B.; Avetyan, D.; Hovhannisyan, L.; Mkrtchyan, G. Genetics of Posttraumatic Stress Disorder—Candidate Genes and Their Implication in the Disease-Associated Molecular Pathomechanisms. In A Fresh Look at Anxiety Disorders; IntechOpen: London, UK, 2015; pp. 65–88. [Google Scholar]
- Ressler, K.J.; Mercer, K.B.; Bradley, B.; Jovanovic, T.; Mahan, A.; Kerley, K.; Norrholm, S.D.; Kilaru, V.; Smith, A.K.; Myers, A.J.; et al. Post-traumatic stress disorder is associated with PACAP and the PAC1 receptor. Nature 2011, 470, 492–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blokland, G.A.M.; McMahon, K.L.; Hoffman, J.; Zhu, G.; Meredith, M.; Martin, N.G.; Thompson, P.M.; de Zubicaray, G.I.; Wright, M.J. Quantifying the heritability of task-related brain activation and performance during the N-back working memory task: A twin fMRI study. Biol. Psychol. 2008, 79, 70–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glahn, D.C.; Winkler, A.M.; Kochunov, P.; Almasy, L.; Duggirala, R.; Carless, M.A.; Curran, J.C.; Olvera, R.L.; Laird, A.R.; Smith, S.M.; et al. Genetic control over the resting brain. Proc. Natl. Acad. Sci. USA 2010, 107, 1223–1228. [Google Scholar] [CrossRef] [Green Version]
- Thompson, P.M.; Martin, N.G.; Wright, M.J. Imaging genomics. Curr. Opin. Neurol. 2010, 23, 368–373. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (s) | Risk Allele/Polymorphism | Subjects | Method Used | Affected Brain Regions | Findings | References |
|---|---|---|---|---|---|---|
| FKBP5 | rs1360780 | Healthy adults and risk allele carriers for PTSD | fMRI (Attention bias task) | Hippocampus | T risk allele carriers showed increased hippocampal activation and altered hippocampal shape | [4] |
| rs1360780 | Traumatized females | DTI, FA | Entorhinal cortex | Lower FA in the left posterior cingulum of risk allele carriers | [5] | |
| COMT | Val158Met | PTSD individuals and healthy adults | MRI | ACC | PTSD-positive participants that were Met carriers showed increased right ACC volume | [6] |
| BDNF | Val66Met polymorphism (rs6265) | Psychologically traumatized disaster survivors and healthy controls | MRI | Prefrontal cortex | Increased DLPFC thickness in the trauma-exposed individuals | [7] |
| COBL | rs406001 | PTSD individuals | sMRI | Prefrontal cortex, amygdala | Alterations in white matter integrity in brain regions associated with emotional processing in affected individuals | [8] |
| SLC6A4 | rs16965628 5-HTTLPR | Post-9/11 veterans with PTSD and trauma-exposed controls | fMRI (working memory task) | Amygdala | rs16965628 SNP modulated task-related ventrolateral PFC activation in patients with PTSD. 5-HTTLPR modulated left amygdala activation during the working memory delay period in S allele carriers with PTSD | [9] |
| ADCYAP1R1 | rs2267735 | Highly traumatized cohort of women | fMRI (threat-processing task) | Amygdala and hippocampus | Increased responses to fearful stimuli in the amygdala and hippocampus of risk allele carriers (CC) | [10] |
| OPRL1 | rs6010719 | Highly traumatized males and females | fMRI (fear-processing task) | Amygdala | risk allele carriers (GG/GC) showed increased functional connectivity between amygdala and posterior insula as compared to the CC genotype | [11] |
| SKA2 | rs7208505 | Trauma-exposed veterans | sMRI | Prefrontal cortex | SKA2 methylation associated with reduced cortical thickness in prefrontal cortex in trauma-exposed veterans | [12] |
| NR3C1 | Mothers with interpersonal trauma | fMRI (mother-child interaction sequences of free-play and separation) | Prefrontal cortex | maternal mPFC activity correlated positively to NR3C1 methylation and negatively to parenting stress and maternal IPV-PTSD in response to a video showing mother and child separation | [13] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nisar, S.; Bhat, A.A.; Hashem, S.; Syed, N.; Yadav, S.K.; Uddin, S.; Fakhro, K.; Bagga, P.; Thompson, P.; Reddy, R.; et al. Genetic and Neuroimaging Approaches to Understanding Post-Traumatic Stress Disorder. Int. J. Mol. Sci. 2020, 21, 4503. https://doi.org/10.3390/ijms21124503
Nisar S, Bhat AA, Hashem S, Syed N, Yadav SK, Uddin S, Fakhro K, Bagga P, Thompson P, Reddy R, et al. Genetic and Neuroimaging Approaches to Understanding Post-Traumatic Stress Disorder. International Journal of Molecular Sciences. 2020; 21(12):4503. https://doi.org/10.3390/ijms21124503
Chicago/Turabian StyleNisar, Sabah, Ajaz A. Bhat, Sheema Hashem, Najeeb Syed, Santosh K. Yadav, Shahab Uddin, Khalid Fakhro, Puneet Bagga, Paul Thompson, Ravinder Reddy, and et al. 2020. "Genetic and Neuroimaging Approaches to Understanding Post-Traumatic Stress Disorder" International Journal of Molecular Sciences 21, no. 12: 4503. https://doi.org/10.3390/ijms21124503