1. Introduction
Cimicifuga racemosa (L.) Nutt. (Ranunculaceae) (synonym of the accepted name
Actaea racemosa L.) extracts have obtained a “well-established use” status in the treatment of menopausal complaints in the Committee of Herbal Medicinal Products conclusions included in the Monograph “Black Cohosh” of the European Medicines Agency (EMA/265439/2018). Although
Cimicifuga racemosa (
C. racemosa) extracts have been widely used for decades [
1,
2,
3], their mechanisms of action remain to be elucidated. Climacteric symptoms include sweating, insomnia, mood changes, headache, vaginal dryness, and, most frequently, hot flushes [
4]. It is well-known that during menopausal transition, the fluctuation of estrogen levels leads to alterations in central nervous system gene expression, neurotransmitter synthesis, and to an impairment of hypothalamic neuronal activity, including mitochondrial dysfunction and imbalanced redox homeostasis [
5]. These alterations promote inflammatory processes and oxidative stress [
6]. The aforementioned changes may largely contribute to the development of hot flushes by narrowing the thermoregulatory set point [
7]. In line with the general view of menopausal complaints being caused by hormonal dysregulation, hormone replacement therapy (HRT) is commonly used to treat menopausal complaints [
6], but has limitations due to adverse side effects, such as headache, uterine bleeding, and enhanced risks for venous thromboembolic disease, breast cancer, stroke, and coronary artery disease [
8,
9].
Menopausal transition and the corresponding symptoms, such as hot flushes, are related to an imbalance in redox homeostasis and are, therefore, often accompanied by the increased formation of reactive oxygen species (ROS) [
10]. We recently reported that the
C. racemosa extract Ze 450 mediates beneficial metabolic effects, protecting against oxidative cell damage in various cell types, including neuronal and liver cells [
11,
12]. Enhanced oxidative stress has been closely linked as a common pathological feature to many age-related disorders, metabolic diseases, and immunological impairments [
13]. Additionally, Ze 450 demonstrated a strong influence on energy metabolism by reducing mitochondrial respiration and promoting glycolysis [
12], similar to the Warburg effect [
14]. To fulfill the energy demand of cells through glycolysis, larger amounts of glucose are required in comparison to the much more efficient oxidative phosphorylation (OXPHOS) in mitochondria.
In line with this, Ze 450 demonstrated antidiabetic effects in an
ob/ob mouse model, as well as beneficial effects on metabolic parameters in ovariectomized rats, similar to the antidiabetic metformin [
15,
16]. Metformin is the first-line treatment option for most patients suffering from diabetes mellitus type II [
17], and extremely interesting insights could be gained by comparing the effects of Ze 450 and metformin on mitochondria. Further, a recent retrospective observational study demonstrated that Ze 450 attenuated weight gain during menopause [
18], confirming that Ze 450 may mediate metabolic effects in patients.
A great deal of research has been performed analyzing the Warburg effect, which describes the metabolic reprogramming towards aerobic glycolysis in cancer cells [
14]. In these cells, such metabolic switches to glycolysis were associated with the activation of key transcriptional regulators like hypoxia-inducible factor α (HIF1α) and cMyc.
Cimicifuga racemosa extract Ze 450 may exert similar regulation of energy metabolism through direct and indirect effects on mitochondrial OXPHOS.
In addition, former studies attributed the mode of action of
C. racemosa extracts to an estrogen-like effect. Notably, this is a controversial discussion in the field, and the majority of more recent reports suggest that
C. racemosa extracts should not be considered as phytoestrogens, and can neither activate estrogen receptors nor mediate estrogen hormone-like effects. In this study, Ze 450 was compared to 17β-estradiol, estrogen receptor agonists (4,4′,4″-(4-propyl-[1H]-pyrazole-1,3,5-triyl)trisphenol (PPT) and diarylpropionitrile (DPN)), and an estrogen receptor modulator (tamoxifen (TM)) to evaluate the similarities and differences of the extract to estrogen-mediated effects in model systems of oxidative stress and on mitochondrial parameters. A well-established in vitro model system of oxidative stress was used [
19] to mimic the imbalance of redox homeostasis during the menopausal transition and emphasize the importance of metabolic changes besides estrogen depletion. Further, effects of Ze 450 and metformin were compared in vivo, in the model organism
Caenorhabditis elegans (
C. elegans) to assess the effects on longevity under basal conditions, and in conditions of oxidative stress exerted by the mitochondrial toxin paraquat.
2. Materials and Methods
2.1. Cimicifuga racemosa Extract Ze 450
The ethanolic (60%
v/v)
C. racemosa dry extract Ze 450 was manufactured from dried roots and rhizomes and obtained from Max Zeller and Soehne AG (Romanshorn, Switzerland), and thus, the indicated doses applied in this study contain 75% native extract. The content of triterpene glycosides was 6.4%. Ze 450 was dissolved in 60% ethanol (
v/v) (Carl Roth GmbH, Karlsruhe, Germany) for all experiments. Ze 450 conforms to the herbal preparation B, which was granted with a well-established use status by a European Union herbal monograph on
C. racemosa of the Herbal Medicinal Product Committee (HMPC) in 2010. The high-performance liquid chromatography (HPLC) fingerprint of the Ze 450 batch used (batch number: 151033) for the current experiments was published [
12].
2.2. Reference Substances
Metformin hydrochloride (Cayman Chemical, Ann Arbor, MI, USA) was dissolved in H2O, 17β-estradiol (Merck KGaA, Darmstadt, Germany) in 60% ethanol and DPN, PPT and 4-OH-Tamoxifen (Merck KGaA, Darmstadt, Germany) were dissolved in DMSO, and all reagents were further diluted into culture medium to the indicated treatment concentrations, unless otherwise stated.
2.3. Cell Culture
HT22 cells (kindly provided by David Schubert, Cellular Neurobiology Laboratory, Salk Institute for Biological Studies, La Jolla, CA, USA) were grown in Dulbecco’s modified Eagle medium (DMEM, Capricorn Scientific GmbH, Ebsdorfergrund, Germany) supplemented with 10% heat-inactivated fetal calf serum (Merck KGaA, Darmstadt, Germany), 100 U/mL penicillin, 100 mg/mL streptomycin (Capricorn Scientific GmbH, Ebsdorfergrund, Germany) and 2 mM L-glutamine (Merck KGaA, Darmstadt, Germany). To induce cell death, erastin (Calbiochem®, Merck KGaA, Darmstadt, Germany) was added to the medium for the indicated amount of time (8–16 h).
2.4. Mitochondrial Isolation
Rat cortical mitochondria were isolated from Sprague-Dawley rats directly after dissection of the prefrontal cortex. All following steps were carried out on ice according to a well-established isolation method [
20]. For cortical tissue, around 50 mg was minced with a scalpel in 1 mL of isolation buffer (300 mM sucrose, 5 mM 2-[[1,3-dihydroxy-2-(hydroxymethyl)propan-2-yl]amino]ethanesulfonic acid (TES), 200 µM ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid (EGTA), 1 mM dithiothreitol (DTT)). Cortical tissue was homogenized by trituration with a 20 G Neoject needle and strained through a 100 µm cell strainer (Life Science, VWR, Radnor, PA, USA). Afterwards, a pump-controlled cell homogenizer (Isobiotec, Heidelberg, Germany) with a constant pump flow rate (700 µL/min) was used to isolate mitochondria from the tissue. The system was primed with 1 mL mitochondrial isolation buffer before usage. Using the cell homogenizer, the cortical suspension was pumped three times with a clearance of 10 µm. Next, the system was rinsed with 1 mL mitochondrial isolation buffer to collect the total homogenate, followed by centrifugation at 800×
g for 10 min at 4 °C (Heraeus™ FRESCO17, Thermo Fisher Scientific, Waltham, MA, USA). The supernatant was transferred to a new 1.5 mL tube and centrifuged at 9000×
g for 10 min at 4 °C. The resulting pellet represents the crude mitochondrial fraction. After resuspension in 1 mL of mitochondrial isolation buffer (MSHE; 70 mM sucrose, 210 mM mannitol, 5 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 1 mM EGTA, 0.5% BSA) a BCA assay for protein determination using the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) was performed. Protein concentration was calculated via linear regression with a BSA standard based on mitochondrial isolation buffer.
2.5. Cell Viability
Cell proliferation was analyzed in real-time by measuring electrical impedance [
21]. Metabolic activity as an indicator of cell viability was quantified using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay [
19]. Viable and metabolically active cells converted MTT (Merck KGaA, Darmstadt, Germany), which was added at a concentration of 2.5 mg/mL for 1 h at 37 °C to the culture medium, into purple formazan. Absorbance was measured at 570 nm vs. 630 nm with FluoStar (BMG Labtech, Ortenberg, Germany) after dissolving in DMSO (Carl Roth GmbH, Karlsruhe, Germany). Cell death of HT22 cells treated with Ze 450, erastin was detected using the Annexin-V-FITC/PI Detection Kit (PromoCell, Heidelberg, Germany) followed by fluorescence-activated cell-sorting (FACS, guava easyCyte, Merck KGaA, Darmstadt, Germany). Annexin-V-FITC was excited at 488 nm, and emission was detected through a 525 ± 30 nm bandpass filter. Propidium iodide (PI) was excited at 488 nm, and fluorescence emission was detected using a 690 ± 50 nm bandpass filter. Data were collected from at least 5000 cells with at least three replicates per condition.
2.6. Lipid Peroxidation
After the indicated treatments, HT22 cells were stained with BODIPY 581/591 C11 (Invitrogen, Carlsbad, CA, USA) for 1 h (37 °C, 4.5% CO2) and harvested for FACS analysis. Lipid peroxidation was analyzed by recording green (emission: 525 nm/30) and red (emission: 585 nm/50) fluorescence with a 488 nm excitation wavelength of at least 5000 cells of at least three replicates per condition. Levels of lipid peroxidation were calculated by the analysis of the shift from green to red fluorescence.
2.7. Mitochondrial ROS Formation
MitoSOX red (Invitrogen, Carlsbad, CA, USA) is selectively targeted to the mitochondria, where it is oxidized by superoxides exhibiting red fluorescence. For detection of mitochondrial ROS formation, MitoSOX red was applied for 30 min at 37 °C and cells were harvested for FACS analysis. Increasing red fluorescence correlating with the formation of mitochondrial ROS was detected by FACS analysis (excitation 488 nm, emission 690 nm/50). Data were collected from at least 5000 cells and three replicates per condition.
2.8. Mitochondrial Membrane Potential
After treatment with Ze 450 and erastin, cells were stained with tetramethylrhodamine, ethyl ester (TMRE) (0.4 nM for 30 min at 37 °C, MitoPT ΔΨm Kit, ImmunoChemistry Technologies, Bloomington, MN, USA) and harvested for TMRE fluorescence measurement via FACS analysis. Upon loss of the mitochondrial membrane integrity and, thus, membrane potential, a loss of TMRE fluorescence can be detected by FACS analysis (excitation 488 nm, emission 690 nm/50). Data were collected from at least 5000 cells and three wells per condition.
2.9. ATP Measurements
ATP levels were detected using the ViaLight™plus Kit (Lonza, Basel, Switzerland). After 24 h post-seeding in 96-well plates (6000 cells per well), cells were treated with Ze 450 and erastin. At the indicated time points after treatment, cells were transferred into a white 96-well plate and ATP levels were analyzed by luminescence detection with FluoStar OPTIMA (BMG Labtech, Ortenberg, Germany).
2.10. Seahorse Measurements on HT22 Cells
To determine the oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) as measures of mitochondrial respiration and glycolysis, respectively, a simultaneous real-time measurement was performed using the XF Extracellular Flux Analyzer (Agilent Technologies, Santa Clara, CA, USA) as previously described [
19]. Briefly, HT22 cells were plated in XF96-well microplates (6000 cells per well, Seahorse Bioscience, Agilent Technologies, Santa Clara, CA, USA) and treated with Ze 450 and erastin. At indicated treatment conditions, the growth medium was replaced by ~180 μL of assay medium (with 4.5 g/L glucose as the sugar source, 2 mM glutamine, 1 mM pyruvate, pH 7.35) and cells were incubated at 37 °C for 60 min. Three to six baseline measurements were recorded before adding the compounds. Oligomycin (ATP synthase inhibitor) (Merck KGaA, Darmstadt, Germany) was injected into port A (20 µL) at a final concentration of 3 µM, Carbonyl cyanide-4-(trifluoromethoxy)phenylhydrazone (FCCP) (uncoupling agent) (22.5 µL into port B) (Merck KGaA, Darmstadt, Germany) at a concentration of 0.5 µM, rotenone/antimycin A (complex I/III inhibitors) (25 µL into port C) (Merck KGaA, Darmstadt, Germany) at a concentration of 100 nM, and 1 µM 2-deoxyglucose (Carl Roth GmbH, Karlsruhe, Germany) (glycolysis inhibitor) at a concentration of 50 mM (27.5 µL into port D), respectively. Three measurements were performed after the addition of each compound (4 min mixing followed by 3 min detection).
2.11. Seahorse Measurement on Isolated Primary Cortical Mitochondria
2.11.1. Electron Flow Assay
Electron flow assay was used to analyze the activity of mitochondrial complex I measuring the OCR with a Seahorse XFe69 Analyzer (Agilent Technologies, Santa Clara, CA, USA). After isolation of cortical mitochondria following established protocols [
20], 10 µg of mitochondrial protein per well were re-suspended in 25 µL/well of mitochondrial assay solution (MAS; 70 mM sucrose, 220 mM mannitol, 10 mM KH
2PO
4, 5 mM MgCl
2, 2 mM HEPES, 1 mM EGTA, 0.2% (
w/v) BSA) with the addition of pyruvate (10 mM) and malate (2 mM). Subsequently, the plate was centrifuged at 2000× g for 20 min at 4 °C (Heraeus™ Megafuge 40R, Thermo Fisher Scientific, Hanau, Germany) to ensure adherence of the mitochondria to the bottom. Afterwards, 155 µL/well MAS was added to the final volume of 180 µL/well. Rotenone, succinate, and antimycin A were diluted in MAS without sucrose and mannitol, while ascorbate and N,N,N’,N’-tetramethyl-p-phenylenediamine (TMPD) were diluted in MAS and serially injected as follows: 2 µM rotenone (port A), 10 mM succinate (port B), 4 μM antimycin A (port C), 10 mM/100 μM ascorbate and TMPD (port D). For investigating complex IV-driven respiration, 1.0 M ascorbate in H
2O (pH 7.2) and 10 mM TMPD in H
2O (pH 7.2) were mixed with an equimolar concentration (10 mM) ascorbate to ensure TMPD remained reduced. One to three baseline measurements were recorded before adding the compounds, and one to three measurements were performed after the addition of each compound (3 min mixing followed by 3 min detection). Data analysis and visualization were performed using XFe Wave software 2.6.1 (Agilent Technologies, Santa Clara, CA, USA) and GraphPad Prism software 6.05 (Agilent Technologies, Santa Clara, CA, USA) displayed as point-to-point oxygen consumption rates.
2.11.2. Coupling Assay
The coupling assay was used to measure the activity of mitochondrial complex II and III recording the OCR with a Seahorse XFe69 Analyzer (Agilent, Santa Clara, CA, USA). The coupling assay was performed as previously described with minor modifications [
22]. Briefly, mitochondria were isolated as described before [
23]. Briefly, 10 µg of cortical and 12 µg of liver mitochondrial protein per well were re-suspended in MAS with the addition of the complex II substrate succinate (10 mM) and the complex I inhibitor rotenone (2 μM). Subsequently, the plate was centrifuged at 2000×
g for 20 min at 4 °C (Heraeus™ Megafuge 40R, Thermo Fisher Scientific, Hanau, Germany) to ensure adherence of the mitochondria to the bottom of the well. Afterwards, 155 µL/well MAS was added to the final volume of 180 µL/well. Modulators of the respiratory chain were diluted in MAS without sucrose and mannitol and serially injected as follows: 4 mM ADP (port A), 2.5 μg/mL oligomycin (port B), 4 μM FCCP (port C), 4 μM antimycin A (port D). One to three baseline measurements were recorded before adding the compounds, and one to three measurements were performed after the addition of each compound (3 min mixing followed by 3 min detection). Data analysis and visualization were performed using XFe Wave software 2.6.1 (Agilent Technologies, Santa Clara, CA, USA), and GraphPad Prism software 6.05 (GraphPad Software Inc., La Jolla, CA, USA), and displayed as point-to-point oxygen consumption rates.
2.12. Rhodamine Measurements
Rhodamine measurement was used to assess mitochondrial membrane potential integrity. Therefore, fluorescence quenching of rhodamine 123 (Thermo Fisher Scientific, Waltham, MA, USA) was measured over 40 min, as described previously [
24]. Mitochondria were plated in a black 96-well plate with 75 µg mitochondrial protein per well and diluted in a buffer containing 200 mM sucrose, 10 mM MOPS-Tris, 5 mM succinate, 1 mM Pi(H
3PO
4), 10 µM EGTA, and 2 µM rotenone (pH 7.3). Loss of mitochondrial membrane potential was induced by FCCP (500 nM), which was injected at the end of the measurement. Fluorescence (excitation: 485 nm/12, emission: 530 nm/10) was measured with a FLUOstar OPTIMA reader (BMG Labtech, Ortenberg, Germany).
2.13. Glutathione Measurement
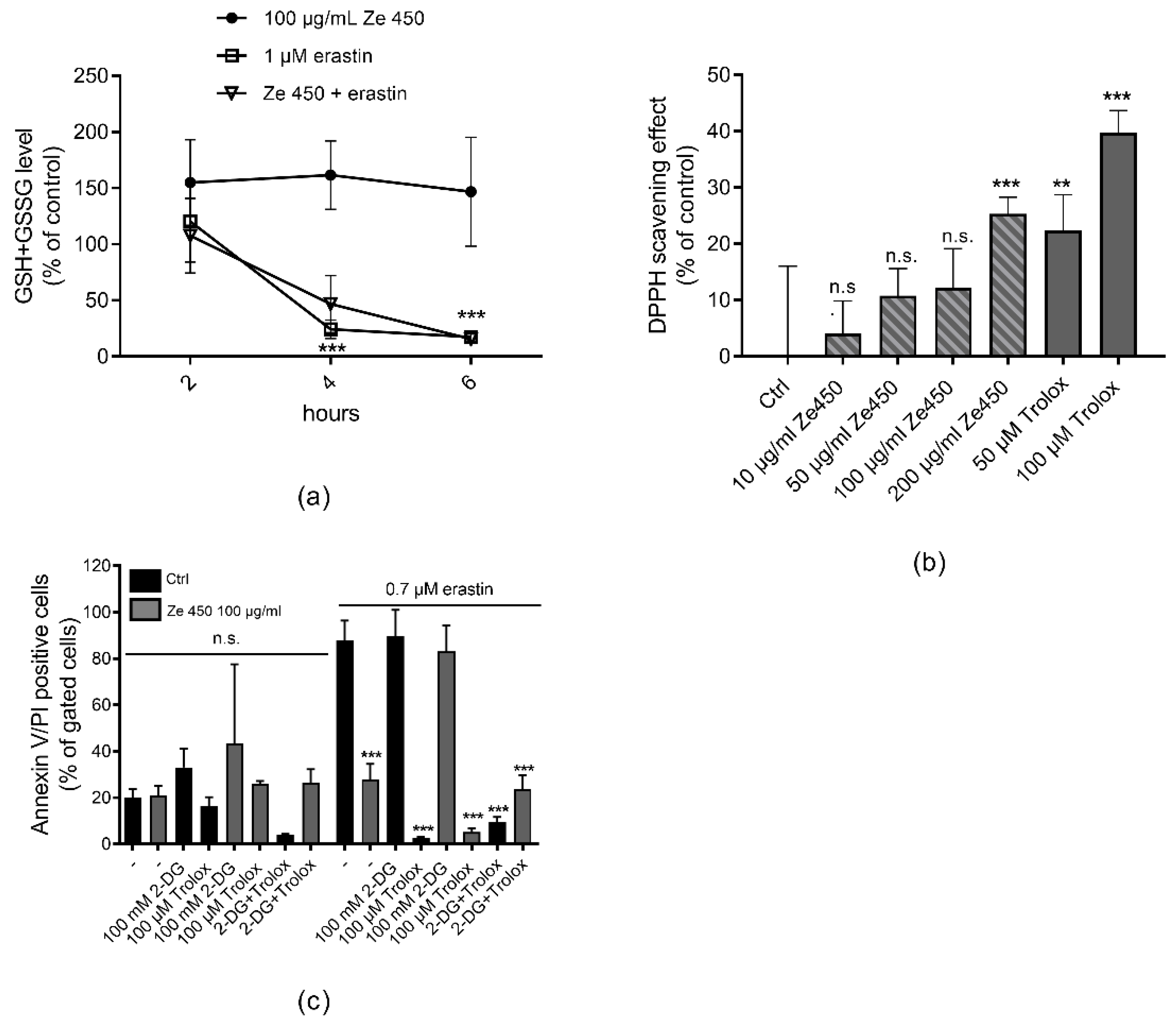
To determine glutathione (GSH) levels, HT-22 cells were seeded in 6-well plates (180,000 cells/well). After treatment with erastin for the indicated amount of time, two to three wells per condition were harvested by scratching, and washed on ice with PBS. GSH measurements were performed using the Glutathione Assay Kit (Cayman Chemical Company, Ann Arbor, MI, USA) following the manufacturer’s protocol. Briefly, cells were re-suspended in MES-buffer (0.4 M 2-(N-morpholino)ethanesulphonic acid, 0.1 M phosphate, 2 mM EDTA, pH 6.0) and homogenized by sonification. Insoluble fragments were removed by centrifugation at 10,000× g for 15 min. The supernatant was deproteinated by the addition of an equal volume of metaphosphoric acid (1.25 M). After incubation for 5 min, the mixture was centrifuged at 17,000× g for 10 min. Subsequently, the supernatant was mixed with a 4 M solution of triethanolamine to increase the pH. After transferral into a 96-well plate, the assay cocktail containing MES-buffer, co-factor mixture, enzyme mixture, and Ellman’s reagent was added. Absorbance was measured at 405 nm after 30 min of incubation. Total GSH amount was determined via standard curve calculation and normalized to the protein content.
2.14. Protein Analysis
Cells were lysed after the indicated time of treatment in 0.25 M D-mannitol, 0.05 M Tris base, 1 mM EDTA, 1 mM EGTA, 1 mM DTT, and 1% TritonX-100 supplemented with protease and phosphatase inhibitor cocktail tablets (Roche Diagnostics, Mannheim, Germany). Liquid nitrogen was used to disrupt cell membranes. Afterwards, the lysate was stored at −80 °C or thawed on ice and centrifuged for 15 min at 10,000×
g at 4 °C. Protein lysate was used from the resulting supernatant. The total protein amount was determined using the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) and 60 µg protein was loaded on a 7.5% polyacrylamide gel. After electrophoresis, the proteins were transferred from the gel onto a PVDF membrane (Roche Diagnostics, Basel, Switzerland). Then, the membranes were incubated with primary antibodies (overnight, at 4 °C), followed by incubation with the corresponding secondary antibody at room temperature. The membrane was incubated with luminol based HRP-Juice Plus (PJK GmbH, Kleinblittersdorf, Germany) for 2 min for protein detection. Visualization was realized with a ChemiDoc XRS system (Bio-Rad Laboratories Inc., Hercules, CA, USA). For densitometric protein analysis, the Bio Rad Image Lab Software was used. For loading control, a housekeeping protein (Actin, GAPDH, Vinculin) was detected in the same way. The antibodies were used as listed in
Table 1.
2.15. Nematode and Bacterial Strain
C. elegans wild-type strain N2 was obtained from the
Caenorhabditis elegans Genetics Center (University of Minnesota, Minneapolis, MN, USA). Nematodes were maintained on nematode growth medium (NGM) agar plates seeded with
E. coli OP50 at 20 °C according to standard protocols [
25]. For all experiments, synchronous populations were generated through a standard bleaching protocol [
26].
2.16. Preparation of NGM Agar Plates
Three grams of NaCl, 2.5 g Bacto™ Peptone (Becton Dickinson, Heidelberg, Germany) and 17 g Bacto™ Agar (Becton Dickinson, Franklin Lakes, NJ, USA) were dissolved in 975 mL distilled water. After autoclaving, 1 mL cholesterol in ethanol (5 mg/mL), 1 mL 1 M CaCl2, 1 mL 1 M MgSO4, and 25 mL 1 M potassium phosphate buffer (pH 6.0) (108.3 g KH2PO4, 35.6 g K2HPO4, H2O to 1 L) were added in this order.
2.17. Cultivation and Treatment
Synchronous larvae were washed twice in M9 buffer (6 g Na2HPO4, 3 g KH2PO4, 5 g NaCl and 0.25 g MgSO4 × 7 H2O per L), counted and adjusted to 10 larvae per 10 µL. Nematodes were raised in cell culture flasks (Sarstedt, Nümbrecht, Germany) or OP50 spread NGM plates. OP50-NGM was added as a standardized food source with a volume 4.4-fold of the larvae containing M9 solution used. L1 larvae were maintained under shaking at 20 °C, reaching adulthood within 3 days. After reaching young adulthood, 48 h prior to the experiment, nematodes were treated with Ze 450 (100, 500, 1000 µg/mL) dissolved in EtOH 1% and metformin (25, 50, and 100 mM), or paraquat (5 mM; Merck KGaA, Darmstadt, Germany) dissolved in M9. Standard OP50, M9, and EtOH 1% served as controls.
2.18. Lifespan Assay
To determine the nematodes lifespan at 20 °C, a modified protocol from Amrit et al. [
27] was applied and synchronized larvae, obtained from egg preparation as stated above, were raised on NGM agar plates spread with standard OP50
E. coli culture. After completing the L4 larval stage, 40 healthy animals per group were transferred to fresh NGM
E. coli containing plates with a sterilized platinum wire. Effectors were incorporated into the OP50 culture with the concentration as needed. Nematodes were transferred to new plates every two days to distinguish between offspring until egg-laying stopped. In line with the separation from eggs and larvae, nematodes were checked for vital signs using a hot platinum wire held next to the animal’s heads. Worms showing no reaction to the heat stimulus were considered dead. The lifespan curves were statistically compared using the log-rank test.
2.19. Heat-Shock Survival Assay
Approximately 10 nematodes were raised per well in a 96-well microplate as mentioned above. After 48 h of incubation with effectors, time till death was determined using a microplate thermo-tolerance assay [
28]. In brief, nematodes were washed off the wells with M9-buffer into 15 mL tubes followed by three additional washing steps. Each well of a black 384-well low-volume microtiter plate (Greiner Bio-One, Frickenhausen, Germany) was prefilled with 6.5 μL M9-buffer/Tween 20 (1%
v/v). Subsequently, one nematode was immersed in 1 μL M9 buffer under a stereomicroscope (Breukhoven Microscope Systems, Capelle aan den Ijssel, The Netherlands). A volume of 7.5 μL SYTOX green (final concentration 1 μM; Life Technologies, Darmstadt, Germany), which penetrates only into cells with a compromised plasma membrane and gets fluorescent after binding to DNA, was added for fluorescent detection. To prevent water evaporation, the plates were sealed with a Rotilab sealing film (Greiner Bio-One, Frickenhausen, Germany). Heat shock (37 °C) was applied and fluorescence measured with a ClarioStar platereader (BMG Labtech, Ortenberg, Germany) every 30 min over the course of 17 h. The excitation wavelength was set at 485 nm, and the emission detected at 538 nm.
2.20. Chemotaxis Assay
Chemotaxis was assessed using a previously published method [
29]. Briefly, agar plates were divided into four quadrants. Sodium azide (0.5 M) was mixed in the same parts with ethanol (95%) as control, or diacetyl (0.5%) as an attractant. Two microliters of either control or attractant solution were added to the center of two opposite quadrants with the same distance to the middle of the plate. Nematodes were washed and separated from larvae as stated above, and approximately 150 animals were placed in the center of the plate. After 1 h, each quadrant was counted, and a chemotaxis index calculated ((number of attractant–number of control)/number total).
2.21. Statistical Analysis
For statistical analysis of the survival curves, the log-rank test (Mantel-Cox test) was employed compared to the corresponding control. A p-value lower than 0.05 was considered to be a significant difference between two survival curves. All data are given as mean + or ±standard deviation (SD). Statistical comparison between treatment groups was performed by analysis of variance (two-way ANOVA) followed by Scheffé’s post hoc test, and a p-value < 0.05 was considered to be statistically significant. Calculations were executed with Winstat standard statistical software 2012.1.0.96 (R. Fitch Software, Bad Krozingen, Germany) and visualized using GraphPad Prism software 6.05 (GraphPad Software, San Diego, CA, USA).
4. Discussion
Although
C. racemosa extracts are widely used to treat menopausal complaints, the exact mechanism of action has not been clarified. The physiological process of menopausal transition is accompanied not only by estrogen depletion but also by age-related metabolic alterations, including for example an increase in body weight. Consequently, meno-pausal women are exposed to enhanced risk factors for metabolic diseases, such as diabetes mellitus type II or osteoporosis [
10,
36]. Ze 450 was recently investigated for its impact on metabolic parameters of menopausal women [
18]. The authors demonstrated that Ze 450 attenuated weight gain in menopausal women. Additionally, it was shown that Ze 450 mediated beneficial effects, including reduction in body weight and improved glucose metabolism in a mouse model of diabetes [
16]. In the present study, we revealed that the
C. racemosa extract Ze 450 mediated advantageous effects on cellular energy metabolism and according to protective effects against oxidative cell damage that were independent of estrogen signaling. In this context, it has been shown that
C. racemosa extract did not affect hormone levels in peri- and postmenopausal women [
37].
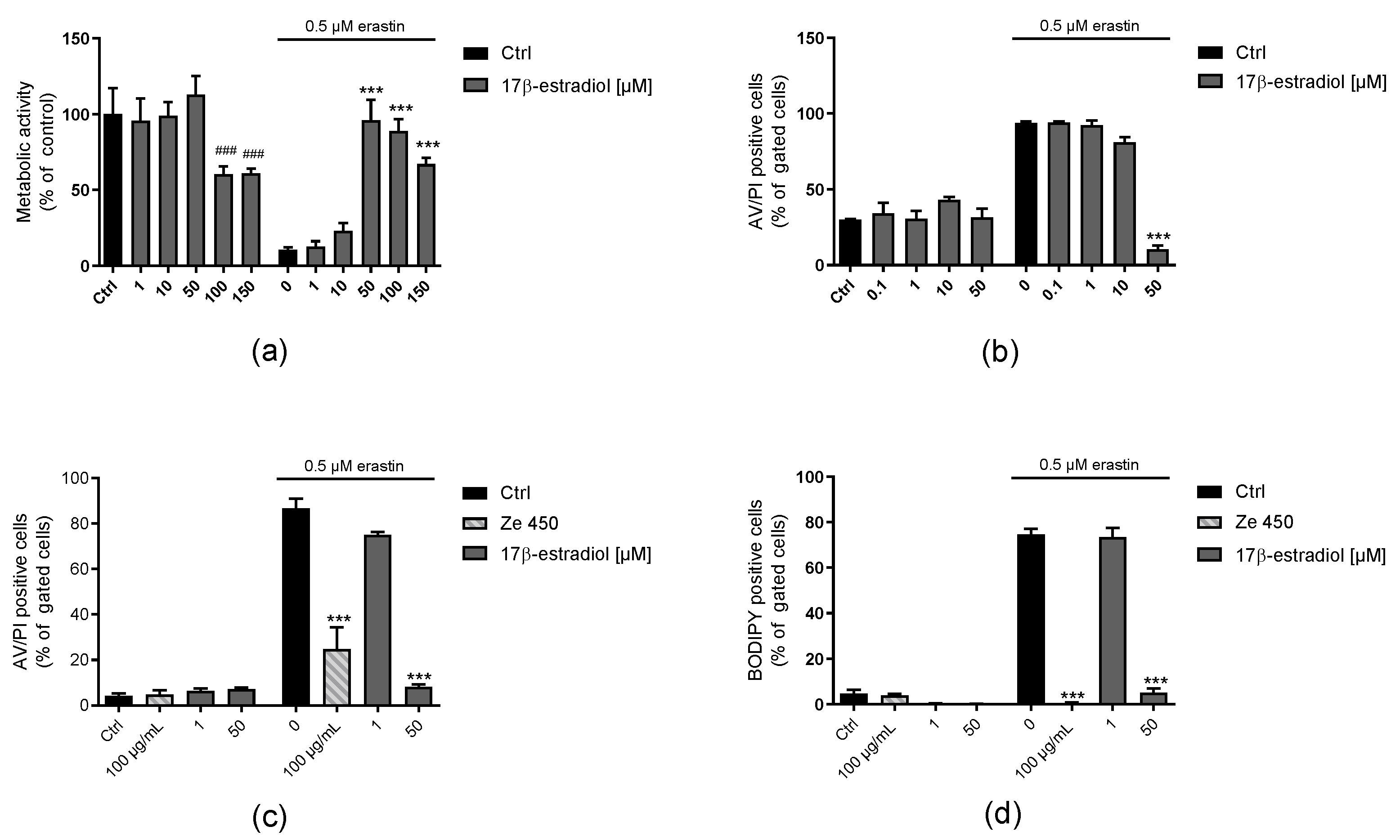
Menopausal complaints are frequently treated with HRT, although this has some limitations due to adverse side effects, such as coronary heart disease, stroke, or increased risk of developing breast cancer. In this study, it was demonstrated that the mechanism of action of Ze 450 is distinct from the effects of 17β-estradiol, estrogen receptor agonists DPN and PPT, and of the SERM tamoxifen (
Figure 4, supporting
Figures S5 and S6). We found that 17β-estradiol protected against erastin-mediated damage, but this effect was attributed rather to its antioxidant properties [
38,
39,
40]. Additionally, mitochondrial respiration was slightly increased by 1 µM of 17β-estradiol compared to control conditions (
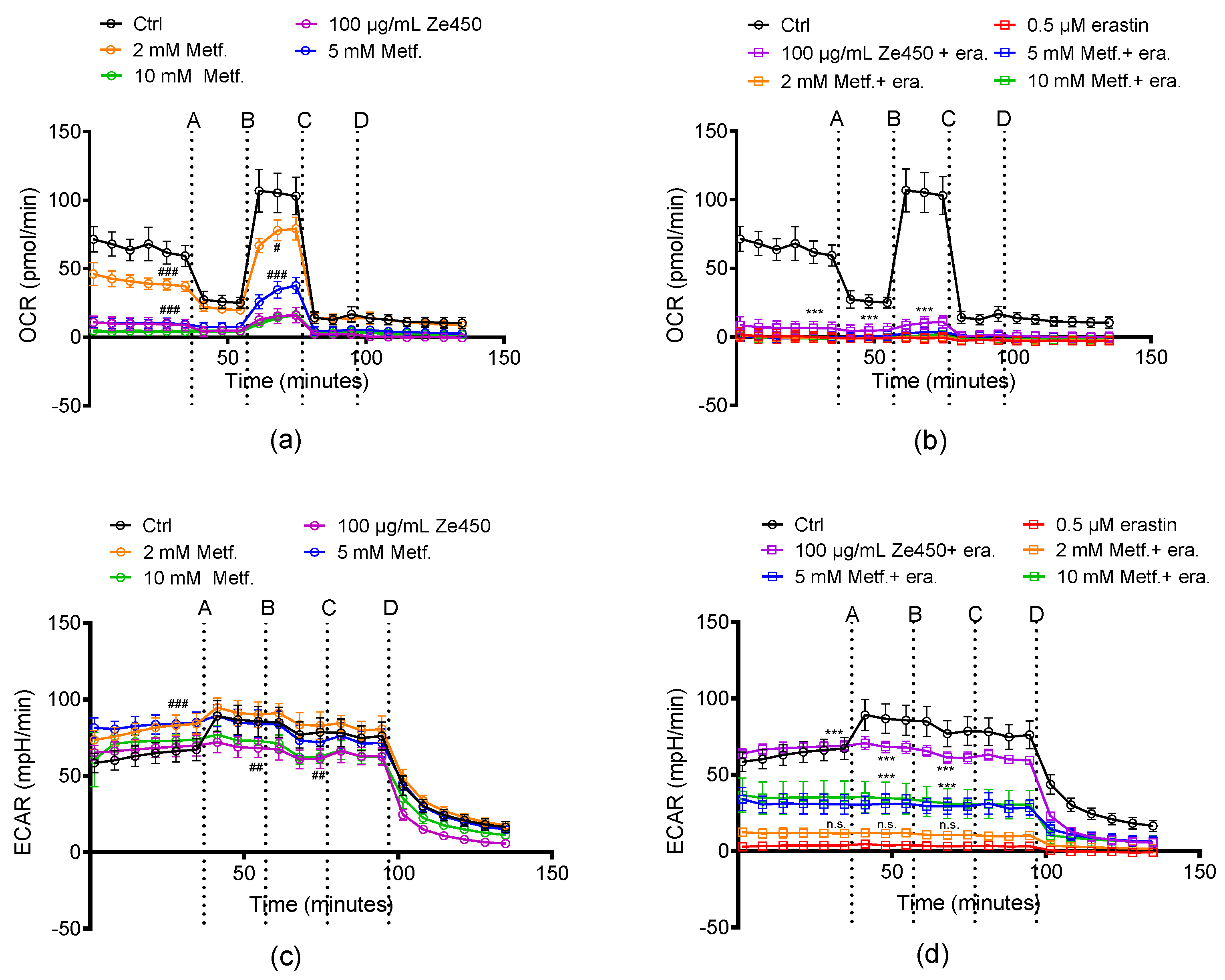
Figure S5). This is in sharp contrast to the effects of Ze 450, which decreased OCR as a mechanistic basis of the observed protective effects.
Moreover, anti-proliferative effects have been demonstrated in vitro [
12], and
C. race-mosa extract has been proposed to mediate protective effects in cancer patients [
41,
42,
43]. Taken together, the findings obtained in the current study indicate a mechanism of action of Ze 450, which is distinct from estrogen-mediated signaling, underlining the importance of
C. racemosa extracts as hormone-free alternatives to treat menopausal complaints, and further suggesting a beneficial influence on metabolic disorders [
15,
16].
Our previous studies [
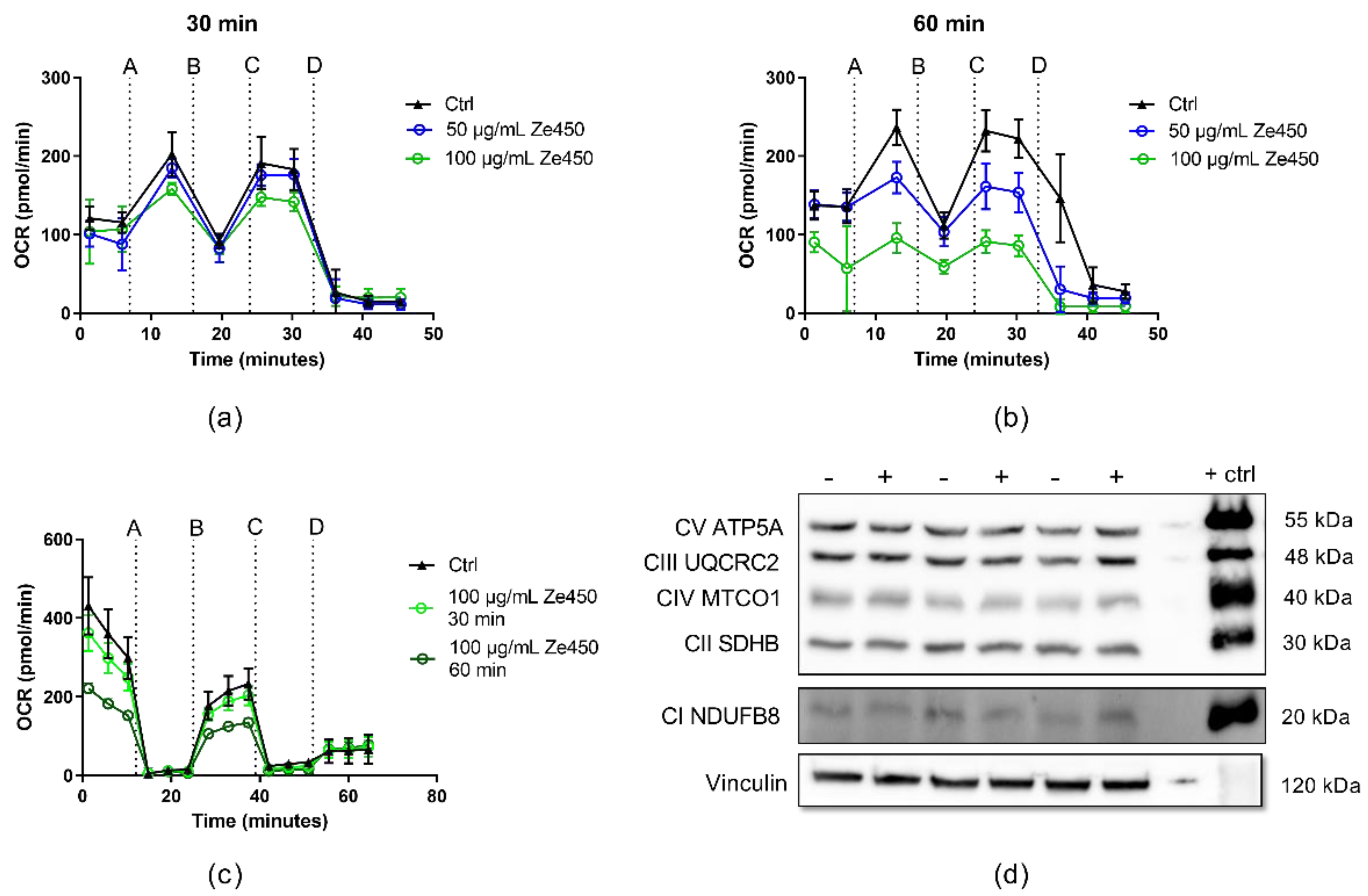
12] and the current results demonstrated that Ze 450 reduced mitochondrial respiration in neuronal cells and promoted an inhibitory effect on mitochondrial complexes in a concentration- and time-dependent manner. Such inhibition of mitochondrial respiration due to reduced mitochondrial complex functionality leads to reduced superoxide production; thus, supporting the hypothesis that Ze 450 mediates protective effects by reducing mitochondrial ROS formation. Consequently, the major production sites of superoxides in mitochondria are inhibited by Ze 450, which subsequently reduces overall oxidative damage to the cells. Our current results point at partial inhibition of CI by Ze 450, but also direct or indirect inhibition of other respiratory complex activities may be involved. So far, we could not detect inhibition of CII or CIII by Ze 450, and, therefore, focused on other mechanisms that may cause the observed metabolic shift from OXPHOS to glycolysis.
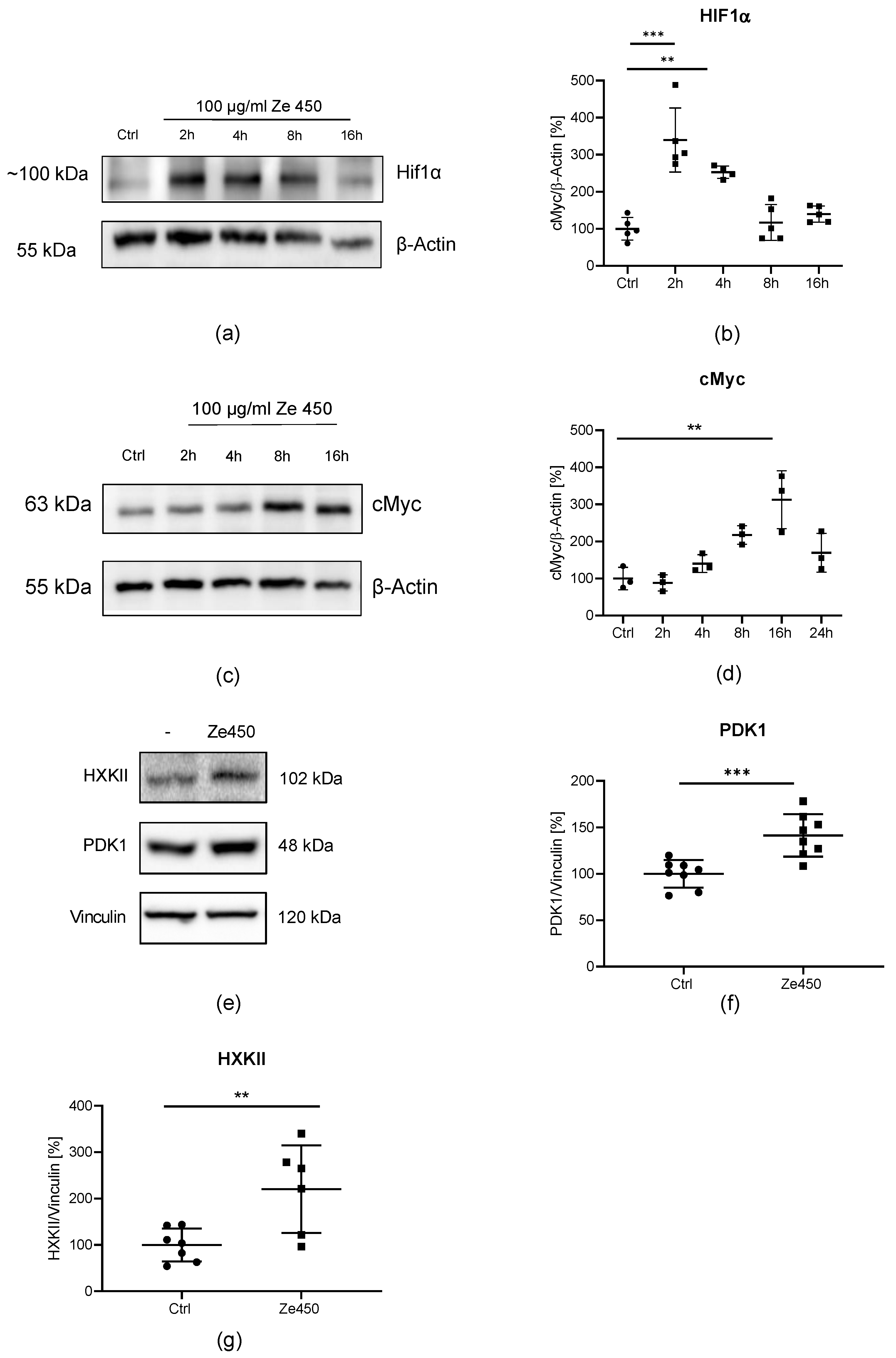
A similar glycolytic switch detected here in neuronal cells exposed to Ze 450 was first described as a fundamental mechanism in cancer cells, and was termed as the Warburg effect. The underlying molecular mechanisms of the Warburg effect are not fully understood, but HIF1α and cMyc have been recognized as master regulators of important glycolytic enzymes like HXKII and PDK1, promoting metabolic reprogramming. In the present model system in neuronal cells, Ze 450 regulated HIF1α and cMyc protein levels in a shifted time dependency, which finally led to an enhanced glycolytic metabolism, by increasing HXKII and PDK1 protein levels.
HIF has been described to be involved in TCA regulation by enhancing PDK1 activity and thereby limiting the entry of pyruvate into the TCA. Consequently, mitochondrial respiration is decreased [
44,
45]. In order to examine the impact on HIF-mediated regulation of energy metabolism, PDK1 protein levels were measured. Concomitant with HIF up-regulation, Ze 450 increased PDK1 protein levels, and mitochondrial respiration measured through OCR was decreased [
12]. Moreover, our results in isolated mitochondria suggested that this effect on OCR was accelerated by a direct inhibition of mitochondrial respiratory chain complexes by Ze 450.
Further, it was demonstrated that HIF is involved in the alteration of COX4 subunits in response to hypoxia in mammalian cells by restoring the electron transfer to O
2 [
46]. This consequently leads to reduced superoxide formation by CI and CIII. In this regard, the regulation of superoxide formation seems to be a crucial metabolic adaption regulated by HIF [
46,
47], and this effect may also contribute to the observed protective effects by Ze 450.
Furthermore, imbalanced redox homeostasis has been linked to many pathological conditions. For example, hot flushes and increased ROS formation have been linked to the metabolic impairments in diabetic patients [
48]. We showed previously that Ze 450 reduced oxidative stress-mediated mitochondrial impairment and cell death in hypothalamic cells [
12] thereby, proposing a potential link to the reported metabolic effects of the
C. racemosa extract; however, the mechanism of action of Ze 450 in experimental settings in vivo and in patients is still unknown. Besides the effective treatment of menopausal complaints, it was suggested that Ze 450 has further therapeutic potential in metabolic diseases. For example, Ze 450 mediated antidiabetic effects in an ob/ob mouse model, which were comparable to metformin [
16], highlighting the potential for Ze 450 as a possible early treatment option for metabolic diseases, such as type II diabetes. This was further supported by clinical data demonstrating reduced weight gain in postmenopausal women taking Ze 450 [
18].
Obesity is one of the most important risk factors for type II diabetes, and the first line of treatment of the disease with the non-insulinotropic antidiabetic drug metformin is known for attenuating body weight gain. Therefore, in the present study, it was of major interest to compare the mode of action of Ze 450 with metformin. Metformin regulates blood glucose levels and fasting insulin levels by reducing gastrointestinal and increasing peripheral glucose uptake [
17,
49,
50]. So far, however, the molecular mechanism of action of metformin requires clarification. It has been suggested that metformin acts at the cellular level as a mitochondrial ETC inhibitor and an activator of AMP-activated protein kinase (AMPK) [
50], thereby interfering with several important regulation pathways of energy metabolism [
51]. Moreover, AMPK activation has been related to neuroprotective effects [
52]. Both metformin and Ze 450 protected against mitochondrial ROS formation, loss of mitochondrial membrane potential and oxidative cell death. Additionally, metformin has been described as a mitochondrial CI inhibitor leading to reduced mitochondrial ROS production [
50,
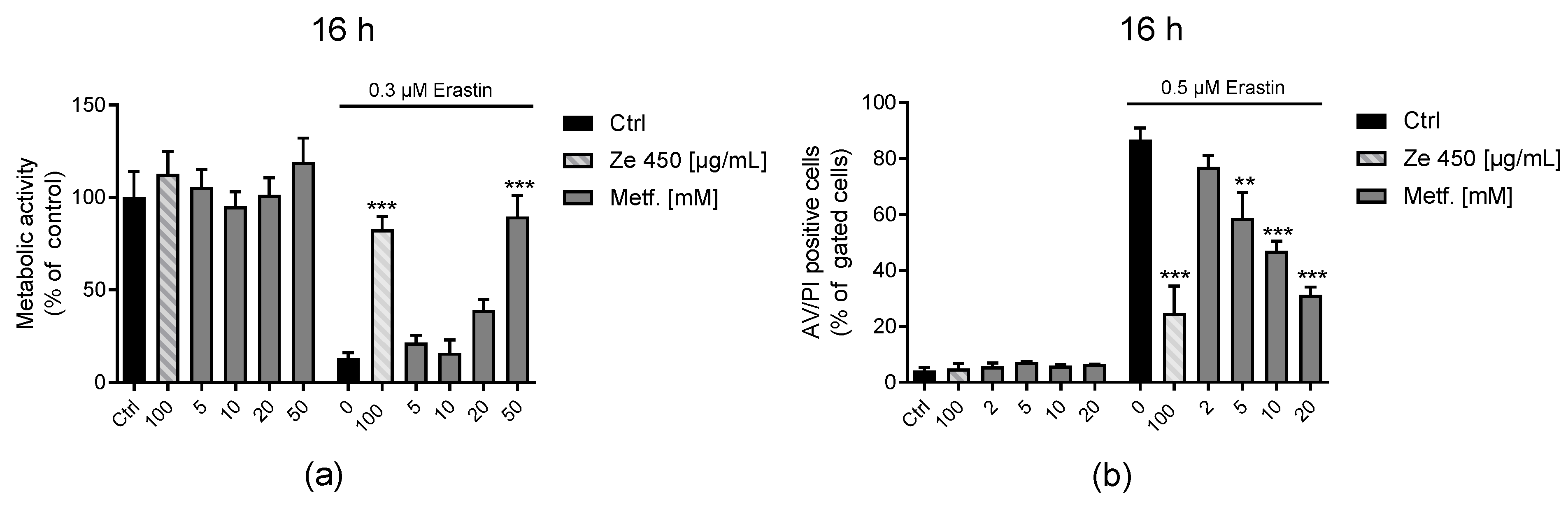
51]. This was also found in the present study, since metformin reduced OCR in a concentration-dependent manner. Here, we found that Ze 450 demonstrated stronger protective effects on the rescue of erastin-mediated ATP depletion compared to metformin. This might be linked to a more pronounced shift towards glycolysis under oxidative stress conditions mediated by Ze 450. These results demonstrate that Ze 450 can interfere with cellular energy metabolism and thereby mediate beneficial effects, which could expand its therapeutic options.
Therefore, it was of major interest to compare the effects of metformin and Ze 450 on a whole organism. For that purpose
C. elegans was used as a suitable in vivo system to evaluate effects on mitochondrial related oxidative stress and longevity [
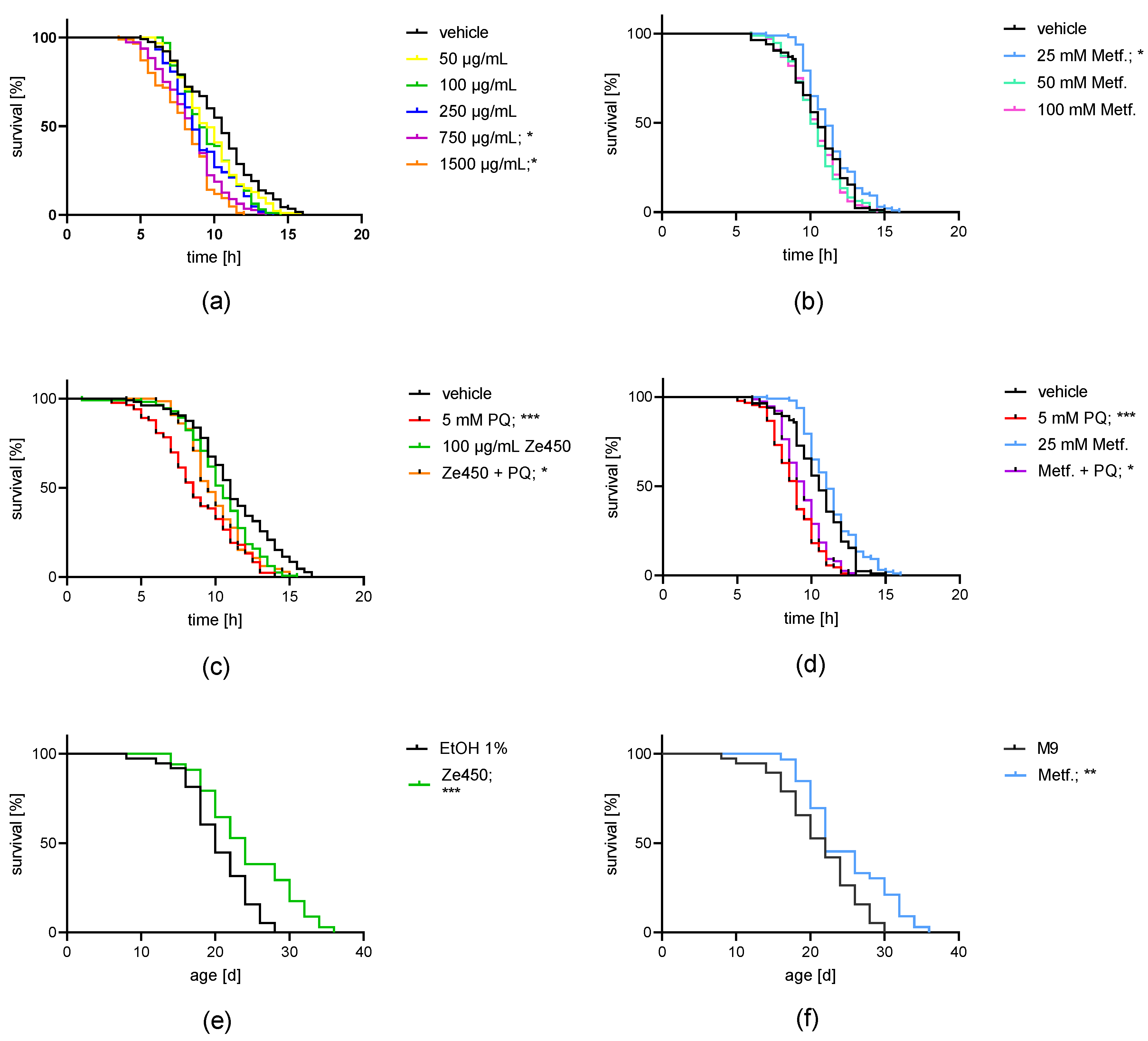
53]. In the present study, 100 μg/mL of Ze 450 protected
C. elegans against PQ-mediated toxicity. Differences between metformin and Ze 450 were identified, since 25 mM metformin enhanced survival in the heat-stress resistance test, while 100 μg/mL of Ze 450 did not. In previous studies, the lifespan extension in
C. elegans by metformin was attributed to a multitude of effects, for example, the modulation of the microbial folate and methionine metabolism [
35], and by activation of AMPK and lysosomal pathways [
34]. Further, metformin-mediated effects on aging are investigated in ongoing clinical trials [
54]. This study may deepen the knowledge about other therapeutic indications beyond its impact on diabetes mellitus type II [
55]. Similarly, the underlying mechanisms promoting longevity and protection against oxidative challenge by
C. racemosa extract in vivo warrant further investigation, also in clinical settings.
The results obtained with C. elegans in vivo emphasizes that Ze 450 has beneficial effects on the whole organism and might contribute to healthy aging. Further, Ze 450 demonstrated protective effects against the oxidative challenge by PQ, confirming the previous findings in vitro, which showed that Ze 450 preserved mitochondrial function and integrity in paradigms of ferroptosis.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}