
Alternaria as an Inducer of Allergic Sensitization
by
 , and
, and
Guadalupe Hernandez-Ramirez
1,2, 
Domingo Barber
3,
Jaime Tome-Amat
1,
Maria Garrido-Arandia
1,2 and
Araceli Diaz-Perales
1,2,* 1
Centro de Biotecnología Y Genómica de Plantas (CBGP, UPM-INIA), Universidad Politécnica de Madrid (UPM), Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), 28223 Madrid, Spain
2
Departamento de Biotecnología-Biología Vegetal, Escuela Técnica Superior de Ingeniería Agronómica, Alimentaria y de Biosistemas, Universidad Politécnica de Madrid (UPM), 28040 Madrid, Spain
3
Departamento de Ciencias Médicas Básicas, Facultad de Medicina, Instituto de Medicina Molecular Aplicada (IMMA), Universidad San Pablo CEU, CEU Universities, 28925 Madrid, Spain
*
Author to whom correspondence should be addressed.
J. Fungi 2021, 7(10), 838; https://doi.org/10.3390/jof7100838
Submission received: 2 September 2021
/
Revised: 1 October 2021
/
Accepted: 4 October 2021
/
Published: 7 October 2021
(This article belongs to the Special Issue Alternaria alternata: Should We Still Consider This Fungus as a Minor Brother in the Etiology of Respiratory Allergy?)
Abstract
:Alternaria alternata is a saprophytic mold whose spores are disseminated in warm dry air, the typical weather of the Mediterranean climate region (from 30° to 45°), with a peak during the late summer and early autumn. Alternaria spores are known to be biological contaminants and a potent source of aeroallergens. One consequence of human exposure to Alternaria is an increased risk of developing asthma, with Alt a 1 as its main elicitor and a marker of primary sensitization. Although the action mechanism needs further investigation, a key role of the epithelium in cytokine production, TLR-activated alveolar macrophages and innate lymphoid cells in the adaptive response was demonstrated. Furthermore, sensitization to A. alternata seems to be a trigger for the development of co-sensitization to other allergen sources and may act as an exacerbator of symptoms and an elicitor of food allergies. The prevalence of A. alternata allergy is increasing and has led to expanding research on the role of this fungal species in the induction of IgE-mediated respiratory diseases. Indeed, recent research has allowed new perspectives to be considered in the assessment of exposure and diagnosis of fungi-induced allergies, although more studies are needed for the standardization of immunotherapy formulations.
1. Introduction
Fungal spores are an abundant component of the atmosphere, constituting the largest proportion of aerobiological particles in the environment [1]. However, of more than 100,000 known fungal species, only a few are responsible for causing airway diseases [2], and most of them belong to the genus Alternaria. Alternaria is the most potent sensitizing aeroallergen source [3], with an important impact on agriculture and health. In the case of agriculture, it is responsible for major crop losses as a plant pathogen [4] and, in terms of human health, it is strongly linked to respiratory disorders, such as asthma, rhinosinusitis [5], pneumonitis, skin infections, and bronchopulmonary mycosis [6,7,8]. Asthma is the most severe disease associated with Alternaria [9,10]. It is characterized by impaired lung function and frequent situations that can lead to death [11]. There is ample evidence demonstrating the unequivocal association among sensitization to A. alternata, the severity of asthma, and hospital admissions to intensive care units [10].
In a broad ISAC-based screening (Thermo Fisher, Uppsala, Sweden) in more than 1000 patients included in grass clinical trials in North America, 22% of the patients were sensitized to mold, and most of them recognized Alternaria [12], suggesting that the clinical relevance of this mold is underestimated.
The goal of this review is to detail the current information regarding A. alternata-induced sensitization and immune responses, with a special focus on the role of its major allergen, Alt a 1.
2. Alternaria alternata as a Plant Pathogen
As a plant pathogen, A. alternata causes black spots on many fruits and vegetables, including cereals, oilseeds, tomatoes, cucumbers, oranges, apples, etc., compromising almost 100 plant species [13]. As a saprophyte, it can infect senescing plants, but it also behaves as a latent fungus that can develop during the cold storage of fruits or remain quiescent for weeks until the fruit ripens.
The genus Alternaria is comprised of more than 350 species which may be saprophytic, endophytic, or even pathogenic in nature [14]. This wide range of action is partly due to the ability of Alternaria to survive in low nutrient media and temperatures ranging from −3 to 35 °C [15]. Its presence has been described in countries in the temperate zone of the planet, such as China, Europe, the United States, Australia and New Zealand [16,17], although there are few data from Africa [18].
Alternaria is usually described based on morphology and/or host specificity. Nowadays, molecular phylogenetic studies have provided a new classification system of twenty-seven sections based upon seven nuclear genes with Alt a 1 and plasmatic membrane calmodulin loci among those genetic marker [19].
3. Alternaria alternata in Human Health
The greatest risks in the case of fungal exposure are the development of respiratory diseases and hypersensitivity. Although hundreds of species that belong to the genus Alternaria are described, only two of them are linked to the development of allergies, i.e., A. alternata and A. chartarum (Ulocladium chartarum) (allergen database, http://www.allergen.org/; accessed on 20 July 2021).
The huge variation in the published data about the prevalence of Alternaria alternata among allergic patients means that it is very difficult to estimate the real prevalence of Alternaria sensitization. This uncertainty may be mainly because of the lack of standardization of extracts for diagnosis [20], until very recently.
In any case, in the literature, several studies can be found where Alternata sensitization is studied among the allergic population. Results from the European Community Respiratory Health Survey I (1990–1992) conducted in 37 centers in 17 countries, involving countries with different geographical parallels such as Australia, the United States of America (USA), the United Kingdom (UK) and Iceland, showed a mean prevalence among populations with respiratory allergic diseases of 4.4% sensitization to A. alternata, varying from 0.2% to 14.4% in the different centers [21,22].
Similarly, a multicenter and open-label study conducted by the European Asthma and Allergy Network (GA2LEN) in 14 European countries have estimated that the prevalence of Alternaria sensitization was, on average, around 6.1%, varying from 2% in Finland to 20% in Greece, in 2009 [23]. In Asia, a cross-sectional study in Iran with about 1000 allergic patients is noteworthy. In this study, sensitization was found to be 5.3%, based on skin prick test reactivity [24]. Finally, in China (Guangdong), sensitization to A. alternata among asthmatic children was 14.9%, while among asthma patients it was 44.9% [16].
These results are in line with the results obtained in the German population, analyzing fungal sensitization in patients with respiratory diseases over 20 years (1998–2017). It was shown that, in the second decade of life, the percentage of fungal sensitization doubled as compared with in the first decade of life [25].
Finally, it is worth mentioning that people working in farms and sawmills are at high risk of A. alternata-induced allergic reactions and infections due to the high level of its allergen in these environments. In addition, among people with respiratory allergy, younger individuals are at higher risk of developing sensitization to A. alternata than older people [5].
4. Alternaria alternata Allergens
Taking into consideration the association between Alternaria and asthma severity [5], a complete definition and immune characterization of the allergen repertoire would contribute by increasing our understanding of this mold as a powerful respiratory allergic disease inducer. To date, a total of seventeen proteins are characterized as allergens in A. alternata (http://www.allergen.org/, accessed on 1 October 2021 and http://www.allergome.org/, accessed on 1 October 2021). This list includes proteins restricted to a small number of taxonomically related fungal species and ubiquitous proteins that were conserved throughout the evolutionary process (Table 1). Most of them have homologues in the other three relevant mold genera in allergy: Cladosporium, Penicillium and Aspergillus, with the exceptions of Alt a 1 and Alt a 13, the most relevant allergens.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Alternaria alternata allergens.
| Allergen | Clinical Relevance | Biological Function | MW (kDa) | Protein (UNIPROT) | Reference |
|---|---|---|---|---|---|
| Alt a 1 | Major (95%) | Unknown | 16.4 and 15.3 | P79085 | De Vouge et al., 1998 [26] |
| Alt a 2 | Inconclusive | Unknown | 22.3–25 | O94095 | Kustrzeba-Wójcicka et al., 2014 [27] |
| Alt a 3 | Minor (5%) | Heat shock protein 70 | 70 | P78983 | De Vouge et al., 1998 [26] |
| Alt a 4 | Minor (42%) | Disulfide isomerase | 57 | Q00002 | Achatz et al., 1995 [28] |
| Alt a 5 | Minor (8–14%) | Ribosomal protein P2 | 11 | P42037 | Achatz et al., 1995 [28] |
| Alt a 6 | Minor (22%) | Enolase | 45 | Q9HDT3 | Simon-Nobbe et al., 2000 [29] |
| Alt a 7 | Minor (7%) | Flavodoxin, YCP4 protein | 22 | P42058 | Achatz et al., 1995 [28] |
| Alt a 8 | Minor (41%) | Mannitol dehydrogenase | 29 | P0C0Y4 | Schneider et al., 2006 [30] |
| Alt a 9 | Minor (5%) | Unknown | 43 | - | Kustrzeba-Wójcicka et al., 2014 [27] |
| Alt a 10 | Minor (2%) | Aldehydede hydrogenase | 53 | P42041 | Achatz et al., 1995 [28] |
| Alt a 12 | Minor | Acid ribosomal protein P1 | 11 | P49148 | Achatz et al., 1995 [28] |
| Alt a 13 | Minor (82%) | Glutathione-S-transferase | 26 | Q6R4B4 | Shankar et al., 2006 [31] |
| Alt a 14 | Minor (11.5%) | Manganese superoxide dismutase | 24 | P86254 | Kustrzeba-Wójcicka et al., 2014 [27] |
| Alt a 15 | Minor (10.2%) | Vacuolar serine protease | 58 | A0A0F6N3V8 | Kustrzeba-Wójcicka et al., 2014 [27] |
| Alt a NFT2 | Minor (0.8%) | Nuclear transport factor 2 | 13.7 | Q8NKB7 | Weichel et al., 2003 [32] |
| TCTP | Minor (4%) | Translationally controlled tumor protein | 20–30 | D0MQ50 | Rid R., et al., 2009 [33] |
| Alt a 1 70 kDa | Minor (87%) | Unknown | 70 | - | Olson et al., 1990 [34] |
While more than 90% of patients sensitized to Alternaria had positive skin test to Alt a 1 [35], it was considered the only well-defined major allergen of this source. However, Alt a 13 is suggested to be another major allergen because of eliciting skin reactions in 14 of 17 patients [31]. Alt a 13 is a 26 kDa glutathione-S-transferase whose function is the detoxification of endogenous and xenobiotic compounds by conjugation of these compounds to reduce glutathione. It is recognized as a cross-reactive allergen in fungal extracts [36]. However, there are insufficient studies, and the studies performed to support its classification as a major allergen were carried out with a small number of patients.
Alt a 1 and Its Flavonol Ligand
Alt a 1 (AAM90320.1, NCBI Protein Database) is a small protein mainly detected in mold spores, which is released at the beginning of germination [37]. Although it is included in its own protein family (AA1) with unknown activity, studies on its homologous Alt b 1 in A. brassicicola and its capacity of inhibiting plant proteins related to pathogenesis (PR5) suggest a biological role in pathogenicity [38,39,40,41]. Other studies support that Alt a 1-like proteins play a key role in the development of plant–fungal interactions [42,43].
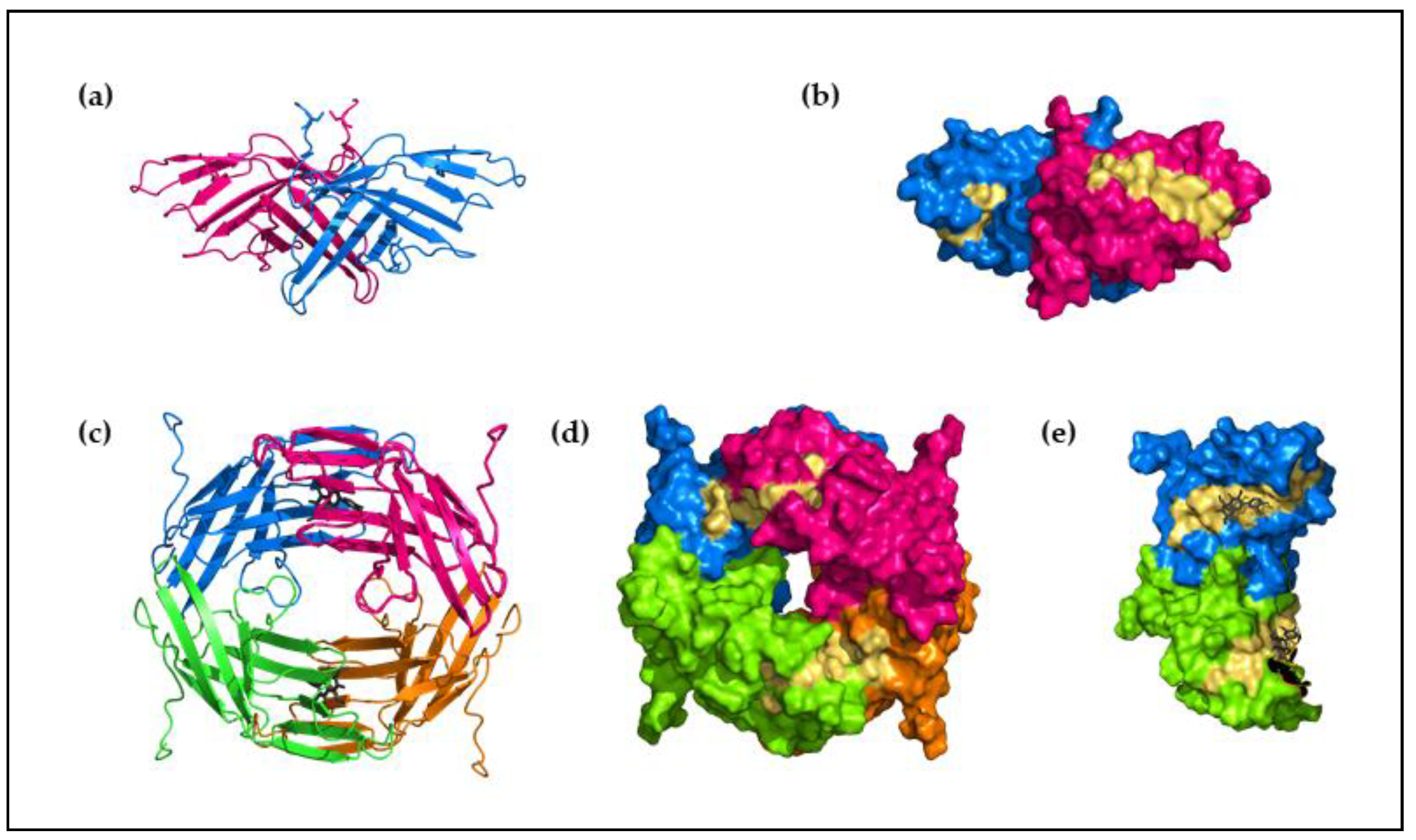
In addition, Alt a 1 is also well characterized as a major allergen and marker of primary sensitization [43]. It is strongly associated with chronic asthma and is declared as a risk factor for the development and exacerbation of asthma. Its high-resolution X-ray crystal structure reveals a unique β-barrel fold formed by eleven β chains that are reported to have no equivalent in the Protein Data Bank [44,45]. The structure is stabilized as a heat-stable, 30 kDa homodimer that dissociates into 15 KDa subunits under reducing conditions or acidic pH. The homodimer shows a “butterfly” shape (Figure 1a) stabilized by an intermolecular disulfide bridge between Cys30 from both monomers and by hydrophobic and polar interactions [44].
In Alt a 1, two linear IgE-binding epitopes (K41-P50 and Y54-K63) are identified [46]; both are localized, exposed on the dimer [47] (Figure 1b), and allow the IgE interaction. In contrast, the tetrameric oligomerization of Alt a 1 (Figure 1c), stabilized by its native ligand, a methyl derivative of quercetin [37,44], hides these residues (Figure 1d), blocking the interaction with the IgE antibodies.
5. Immunological Mechanism of Alternaria Action
Several studies have demonstrated that Alternaria extract is able to provoke allergic responses in mice; the extract is composed of a complex mixture of proteins, mycotoxins, cell wall fragments, chitin, mannans, and B-1,2-glucans [48]. However, there are no data concerning the contribution of each specific component of this mixture in proinflammatory responses. Within this context, the current information about the innate and adaptive immune responses associated with Alternaria action are summarized further with special attention to the role of Alt a 1 in allergy triggering. An overview of the events that take place is described in this section and is shown in Figure 2.
5.1. First Line of Defense: The Early Response
The human respiratory epithelium is a mucosal surface composed of ciliated cells, mucous-producing cells, and undifferentiated basal cells. It is the first site of contact for all inspired substances, acting as both a physical and an immunological barrier to pathogens and external particles to aid the maintenance of normal airway function [49]. Therefore, when mold spores are inhaled and reach the upper airways, the external epithelial barrier responds by producing antimicrobial molecules, proinflammatory cytokines, and chemokines for immune cell recruitment [50]. Many studies on the effects of fungal extracts in the lungs have focused on how they are recognized by pattern recognition receptors (PPRs) and/or toll-like receptors (TLRs) [51,52,53].
Alternaria, by itself, can induce the activation of the epithelium, inducing a quick release of alarmins such as thymic stromal lymphopoietin (TSLP), IL-25, and IL-33, and even the production of other inflammatory cytokines such as IL-6 and IL-8 [54,55,56,57,58,59]. Likewise, other cellular responses are associated with Alternaria, such as ATP release, calcium signaling, calprotectin production, or eosinophil activation [60,61]. Interestingly, some of these studies were conducted using heat inactivated Alternaria extracts, which suggests that protease activities are not necessary to induce asthma pathogenesis by this mold.
Alt a 1 is released in large quantities by spores at pH values around 5.0–6.5, carrying its flavonol and catechol ligands [37]. On the one hand, Alt a 1 interacts with the SLC22A17 receptor in bronchial epithelial cells and mediates the production of IL-33 and IL-25 [62]. On the other hand, this interaction does not result in an increase in barrier permeability, and thus does not explain how Alt a 1 reaches the basolateral side and comes in contact with the immune system cells.
Indeed, Hayes et al. [53] reported that Alt a 1 induced the production of IL-8, MCP-1, and Groa/b/g in airway epithelial cells in a TLR2/4-dependent manner, suggesting that Alt a 1 could be recognized by different subsets of immune cells such as alveolar macrophages. These immune cells, together with the epithelial barrier, are the first line of defense in lungs, by constantly sensing the local environment [63,64]. Therefore, given the well-known function of alveolar macrophages, they could contribute to Alt a 1 incorporation and processing, triggering the production of proinflammatory cytokines. This inflammatory environment, together with the presence of alarmins from epithelial cells, may further impair the epithelial barrier. Moreover, these cells could be implicated in the presentation of Alt a 1 to adaptative immune cells [65].
5.2. Type 2 Immune Response: The Adaptative Response
Asthma and respiratory allergy are two diseases that are still not fully understood, with several unrelated explanations for the mechanisms of how a patient acquires these phenotypes. However, numerous studies have reported a strong correlation between Alternaria inhalation and the development of asthma or allergic rhinitis [66,67,68,69].
A. alternata promotes the production of IL-4, IL-10, and IL-6 in antigen-presenting cells (APCs), together with an increase in the expression of MHC class-II, CD40, CD80, and OX40L, which are costimulatory molecules implicated in type 2 development, in in vitro approximations [70]. Moreover, Alternaria can inhibit the endosome receptor TLR3 and the production of IFN-beta, IP-10, and I-TAC to inhibit defense mechanisms for viral and fungal infections [71].
In the literature, there are many murine models, in which the application of extract or only spores show airway hyperreactivity, mucus hypersecretion, humoral response, lung inflammation, and eosinophilic infiltration, among other effects related to airway diseases. Furthermore, in these models, the production of type 2 cytokines IL-4, -5, and -13, as well as high levels of Alternaria specific IgE is also observed [54,72].
In the last decade, many studies have focused on the critical role of innate lymphoid cells type 2 (ILC2s) in Alternaria-mediated immunopathology in the airways [59,69,73,74]. Tissue resident ILC2s are described as highly responsive, early effectors in type 2 responses, functioning as a prominent source of typical type 2 cytokines IL-5, IL-9, and IL-13, long before an adaptative response is initiated [75]. By studying the role of this cell population in mice exposed to Alternaria, a rapid type 2 cytokine response driven by an activated lung ILC2 population has been established. Therefore, the restoration of lung pathology in mice deprived of ILC2s and exposed to Alternaria has finally confirmed the contribution of ILC2s to allergic airway inflammation in animal models [54,76,77].
6. The Trojan Horse: Alternaria as an Inductor of Other Allergies
Airborne spore counts are often 1000-fold greater than pollen counts, and exposure is often longer in duration. Indeed, there is a clear correlation between high atmospheric concentrations of certain mold spores (for instance during thunderstorms) and an increase in hospital admissions due to asthma exacerbations [80]. In addition, many studies have noted that asthma severity is often more closely associated with Alternaria sensitization than with pollen [81]. Within the clinical relevance of Alternaria, its potential as an indoor mold should not be underestimated, since it can grow in dampness and even on fruits and vegetables without visible signs of infection [41]. Thus, it is reasonable to consider that molds such as Alternaria may be initiators of allergic responses, either food and/or respiratory diseases. The next sections focus on two relevant examples regarding co-sensitization events with A. alternata, which could aid in understanding and predicting sensitization and improving allergy diagnostic methods.
6.1. Alternaria and Grass Pollen Cosensitization
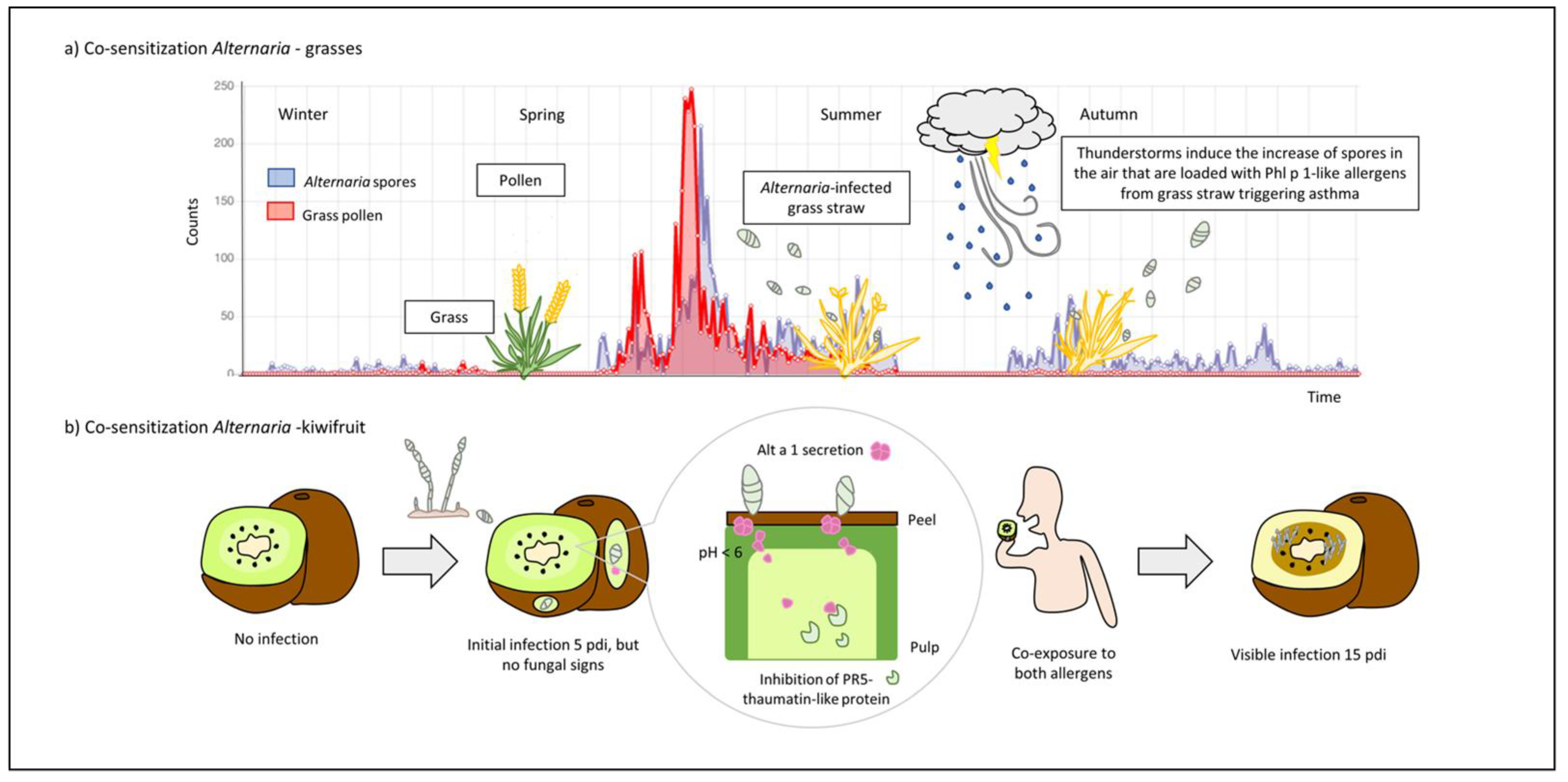
Continuously changing environments lead to differences in exposure and sensitization to allergens, which in turn vary depending on geographic region [80]. For example, the monitoring of Alternaria conidia in 12 stations in Spain showed three different patterns of spore dynamics and concentration levels correlated to daily temperatures and precipitation [82]. Likewise, correlations between Alternaria spore counts and symptoms of rhinitis and/or asthma have been described for Alternaria-sensitized patients [83]. Thus, the development of logistic regression models for predicting the daily concentrations of airborne spores could also aid to predict symptoms or improve the diagnosis of fungi-induced airway diseases [82]. Hernandez-Ramirez et al. [84] described a co-exposure event for grass pollen-allergic patients based on a correlation between high levels of Alternaria spores and asthmatic exacerbations suffered by the patient population when pollen was no longer present. After the identification of Phl p 1-like proteins in grass straw and their ability to bind over the surface of Alternaria spores, the authors suggested that mold spores may act as allergen carriers favoring the development of co-sensitization between Alternaria and other aeroallergens-like group 1 allergens (Figure 3a). It has long been described that the levels of respirable particles, including pollen grains and fungal spores, increased following rainfall events, as well as the release of their allergenic components as a result of the force of storms, moisture conditions, and/or disturbances such as wind and lawn mowing [85]. Interestingly, when the sensitization profile was studied in patients with thunderstorm asthma, the clear co-sensitization to both aeroallergen sources suggested that more than one hit was required [81].
Furthermore, the role of Alternaria as a clinically relevant model for environmental exposure was also studied in asthma developed by ragweed pollen allergy. Consistent with previous reports that showed the adjuvant effects of Alternaria to induce a potent type 2 response in OVA-sensitized mice [86], Kobayashi et al. [87] obtained the same result for mice sensitized to short ragweed pollen, whose humoral responses were minimal when they were exposed to the extract alone. In this study, the authors also introduced the key role of DCs exposed to Alternaria to enhance type 2 polarization of CD4+ T cells. Likewise, a single challenge with Alternaria extract resulted in increased eosinophils, peribronchial inflammation, and mucus production, as well as type 2 cells and ILC2s recruitment to the airway in ryegrass challenged mice, suggesting that the combination of mold spores and increased pollen allergen exposure during thunderstorms may be responsible for severe asthma [57].
6.2. Alternaria and Kiwifruit Cosensitization
In addition to Alternaria effects as airborne mold, Alternaria spp. are also well known to infect a wide variety of fruits and vegetables [4]. Alternaria contamination is also documented to trigger food allergy, as reported by Gómez-Casado et al. [41] In controlled infection assays of kiwifruit with Alternaria spores, the authors demonstrated that Alt a 1 was released from spores to kiwifruit pulp, partially inhibiting the activity of a defense plant protein, the PR5 thaumatin-like protein, when hyphal growth was not yet developed. This study found the origin of a co-sensitization phenomenon between Alt a 1 and Act d 2, a major allergen and PR5 of kiwifruit, caused by the consumption of kiwifruits infected with Alternaria but apparently in good conditions (Figure 3b). According to the early detection of Alt a 1 in infected kiwifruits, similar results were obtained in citrus fruits, highlighting the value of Alt a 1 as a reliable and specific marker of fungal contamination [88].
At the same time, the Alternaria-spinach syndrome was also reported in mold-sensitized patients with food allergic reaction after mushroom (Agaricus bisporus) and spinach ingestions [89,90].
Accordingly, food contamination with fungal spores may be the cause of some adverse reactions that patients suffer from, who apparently were not sensitized in the past, and obtained negative results in previous skin prick tests for the diagnosis of a food allergy.
7. Immunotherapy for Alternaria Sensitization
There is limited evidence on the benefit of allergen immunotherapy (AIT) in Alternaria allergy, the only mold allergy with high prevalence [91]. The European Academy of Allergy and Clinical Immunology (EAACI) does not recommend AIT using mold extracts in children. This opinion was based on a lack of efficacy and safety [92]. On the other hand, The American Academy of Allergy, Asthma and Immunology (AAAAI) has concluded that such an AIT might be effective. An international consensus on immunotherapy indicated the possibility of AIT for mold allergy, but only with standardized extracts [93].
This type of allergy is relatively frequent in children under 10 years of age as a mono-sensitizing allergen [94,95]. In adults, Alternaria sensitization is mostly associated with another allergy, thus making AIT questionable. With current regulations, performing clinical trials in pediatric patients requires previous clinical experimentation in adults. Moreover, in many cases, the asthma associated with Alternaria might be a contraindication, hence the design of appropriate clinical trials is difficult and expensive, a fact that has limited the building of clinical evidence. Traditionally, the active ingredients of any AIT product have consisted of protein mixtures [96], causing, for instance, great variability in the concentration of allergens in the existing products for in vivo allergy diagnosis and AIT [20]. Assuring the correct content of Alt a 1 in AIT preparation is critical to initiate any AIT intervention in patients with Alternaria allergy, as was shown in an interesting therapeutic approach with positive initial results based on the use of purified Alt a 1 in AIT preparation [35].
Many studies have highlighted the effectiveness of AIT for Alternaria in patients with allergic rhinitis and/or bronchial asthma [12,97,98,99,100,101] This applies to children and adults. This is the only mold allergen for which medical documentation has confirmed a significant improvement after treatment in controlled studies. The effectiveness of such AIT is dependent upon the quality of the vaccine. The development of methods for the extraction and assessment of the main allergen, Alt a 1, has occurred because standardization made it possible to extract Alternaria alternata. Studies that used a vaccine containing the standardized Alternaria alternata extract have confirmed the effectiveness of such an AIT [102]. This treatment led to reduced clinical allergic rhinoconjunctivitis symptoms, bronchial asthma, reduced drug used, decreased serum specific IgE levels, and increased serum specific IgG4 levels to Alternaria. It was also important to specify the cumulative Alt a 1 antigen doses that patients received during AIT in these trials, since this was one of the criteria for assessing the effectiveness of AIT. These authors simultaneously observed very good tolerance of such vaccines. Adverse events were observed in less than 1% of all injections. There were mainly local reactions and only a few systemic reactions, which included dyspnea, cough, and rhinitis [103,104,105]. Earlier, an older vaccine (without proper allergen extract standardization) induced many adverse systemic reactions. This greatly limited the possibilities of such treatment [106,107,108].
Despite the effectiveness of AIT treatment with Alternaria, there is a lack of long-term follow up of patients after such treatment. There is no feedback on how long the effect persists after desensitization to Alternaria. Long-term observations of the effectiveness of AIT are difficult and expensive to implement.
8. Conclusions
Fungal allergies are increasing in recent years, and thus it is essential to understand and describe the molecular events associated with them. In addition, symptomatology is often associated with the development of severe hypersensitivity reactions in the airways that compromise the health and quality of life of susceptible individuals.
One of the main fungi responsible for this pathology is A. alternata, and although it is traditionally described as one of the main causes of childhood asthma, its prevalence is also increasing in adults. Because of this increase, a comprehensive understanding of the mechanisms and diagnosis of allergic symptoms caused by the mold are necessary to acknowledge why and how it can induce lung inflammation prior to the development of asthma.
However, despite efforts to describe the potential of Alternaria as an inducer of respiratory allergic inflammation, questions about the individual effects of its allergenic components, especially Alt a 1 as the major allergen, remain unresolved. It is only by means of a complete characterization of immunological mechanisms triggered by this mold that we can improve the diagnosis and, above all, the design of new treatment formulations.
Author Contributions
G.H.-R., J.T.-A., M.G.-A. and A.D.-P. wrote the manuscript. D.B. and A.D.-P. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Government (MINECO, grant BIO2017-84548-R; MICINN, grant PID2020-113629RB-I00) and the Comunidad de Madrid (FOODAL-CM: S2018/BAA-4574). J.T.A. received a grant from the Instituto de Salud Carlos III (ISCIII) co-founded by FEDER Thematic Networks and Cooperative Research in Health, RETICS; ARADyAL (RD16/006/003). G.H.R. was funded by the European Commission with the AllerScreening project (H2020-NMBP-X-KET-2017-768641).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Damialis, A.; Kaimakamis, E.; Konoglou, M.; Akritidis, I.; Traidl-Hoffmann, C.; Gioulekas, D. Estimating the Abundance of Airborne Pollen and Fungal Spores at Variable Elevations Using an Aircraft: How High Can They Fly? Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rick, E.M.; Woolnough, K.; Pashley, C.H.; Wardlaw, A.J. Allergic Fungal Airway Disease. J. Investig. Allergol. Clin. Immunol. 2016, 26, 344–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Ahmad, M.; Jusufovic, E.; Arifhodzic, N.; Rodriguez, T.; Nurkic, J. Association of Molds and Metrological Parameters to Frequency of Severe Asthma Exacerbation. Allergy Asthma Clin. Immunol. 2019, 15, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Adeniji, A.A.; Babalola, O.O.; Loots, D.T. Metabolomic Applications for Understanding Complex Tripartite Plant-Microbes Interactions: Strategies and Perspectives. Biotechnol. Rep. 2020, 25. [Google Scholar] [CrossRef]
- Gabriel, M.F.; Postigo, I.; Tomaz, C.T.; Martínez, J. Alternaria Alternata Allergens: Markers of Exposure, Phylogeny and Risk of Fungi-Induced Respiratory Allergy. Environ. Int. 2016, 89–90, 71–80. [Google Scholar] [CrossRef]
- Chowdhary, A.; Agarwal, K.; Kathuria, S.; Gaur, S.N.; Randhawa, H.S.; Meis, J.F. Allergic Bronchopulmonary Mycosis Due to Fungi Other than Aspergillus: A Global Overview. Crit. Rev. Microbiol. 2013, 40, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Pfavayi, L.T.; Sibanda, E.N.; Mutapi, F. The Pathogenesis of Fungal-Related Diseases and Allergies in the African Population: The State of the Evidence and Knowledge Gaps. Int. Arch. Allergy Immunol. 2020, 181, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Hattab, Z.; Lasfar, N.B.; Abid, M.; Bellazreg, F.; Fathallah, A.; Hachfi, W.; Letaief, A. Alternaria Alternata Infection Causing Rhinosinusitis and Orbital Involvement in an Immunocompetent Patient. New Microbes New Infect. 2019, 32. [Google Scholar] [CrossRef]
- Sachdeva, K.; Do, D.C.; Zhang, Y.; Hu, X.; Chen, J.; Gao, P. Environmental Exposures and Asthma Development: Autophagy, Mitophagy, and Cellular Senescence. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Agnihotri, N.T.; Saltoun, C. Acute Severe Asthma (Status Asthmaticus). Allergy Asthma Proc. 2019, 40, 406–409. [Google Scholar] [CrossRef]
- Fergeson, J.E.; Patel, S.S.; Lockey, R.F. Acute Asthma, Prognosis, and Treatment. J. Allergy Clin. Immunol. 2017, 139, 438–447. [Google Scholar] [CrossRef] [Green Version]
- Nolte, M.; Barber, D.; Maloney, J.; Li, Z.; Kaur, A.; Galan, A.; Andersen, J.S.; Nolte, H. Timothy Specific IgE Levels Are Associated with Efficacy and Safety of Timothy Grass Sublingual Immunotherapy Tablet. Ann. Allergy Asthma Immunol. 2015, 115, 509–515. [Google Scholar] [CrossRef]
- Meena, M.; Gupta, S.K.; Swapnil, P.; Zehra, A.; Dubey, M.K.; Upadhyay, R.S. Alternaria Toxins: Potential Virulence Factors and Genes Related to Pathogenesis. Front. Microbiol. 2017, 1451. [Google Scholar] [CrossRef] [PubMed]
- Dalinova, A.A.; Salimova, D.R.; Berestetskiy, A.O. Fungi of the Genera Alternaria as Producers of Biological Active Compounds and Mycoherbicides. Appl. Biochem. Microbiol. 2020, 56, 256–272. [Google Scholar] [CrossRef]
- Ji, Y.; Hu, W.; Liao, J.; Xiu, Z.; Jiang, A.; Yang, X.; Guan, Y.; Feng, K.; Saren, G. Effect of Ethanol Vapor Treatment on the Growth of Alternaria Alternata and Botrytis Cinerea and Defense-Related Enzymes of Fungi-Inoculated Blueberry During Storage. Front. Microbiol. 2021, 12, 1. [Google Scholar] [CrossRef]
- Guan, K.; Liu, B.; Wang, M.; Li, Z.; Chang, C.; Cui, L.; Wang, R.-Q.; Wen, L.-P.; Leung, P.S.C.; Wei, J.-F.; et al. Principles of Allergen Immunotherapy and Its Clinical Application in China: Contrasts and Comparisons with the USA. Clin. Rev. Allergy Immunol. 2019, 57, 128–143. [Google Scholar] [CrossRef] [PubMed]
- Zureik, M.; Neukirch, C.; Leynaert, B.; Liard, R.; Bousquet, J.; Neukirch, F. Sensitisation to Airborne Moulds and Severity of Asthma: Cross Sectional Study from European Community Respiratory Health Survey. Br. Med. J. 2002, 325, 411–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pefura-Yone, E.W.; Mbele-Onana, C.L.; Balkissou, A.D.; Kenmegne-Noumsi, E.C.; Boulleys-Nana, J.R.; Kolontchang-Yomi, B.L.; Theubo-Kamgang, B.J.; Ndjeutcheu-Moualeu, P.I.; Ebouki, E.R.; Kengne, A.P. Perennial Aeroallergens Sensitisation and Risk of Asthma in African Children and Adolescents: A Case-Control Study. J. Asthma 2015, 52, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.P.; Rotondo, F.; Gannibal, P.B. Biodiversity and Taxonomy of the Pleomorphic Genus Alternaria. Mycol. Prog. 2016, 15, 1–22. [Google Scholar] [CrossRef]
- Esch, R.E.; Codina, R. Fungal Raw Materials Used to Produce Allergen Extracts. Ann. Allergy Asthma Immunol. 2017, 118, 399–405. [Google Scholar] [CrossRef]
- Bousquet, P.J.; Chinn, S.; Janson, C.; Kogevinas, M.; Burney, P.; Jarvis, D. Geographical Variation in the Prevalence of Positive Skin Tests to Environmental Aeroallergens in the European Community Respiratory Health Survey, I. Allergy 2007, 62, 301–309. [Google Scholar] [CrossRef]
- Bousquet, P.J.; Hooper, R.; Kogevinas, M.; Jarvis, D.; Burney, P.; Chinn, S.; Luezynska, C.; Vermeire, P.; Kesteloot, H.; Bousquet, J.; et al. Number of Allergens to Be Tested to Assess Allergenic Sensitization in Epidemiologic Studies: Results of the European Community Respiratory Health Survey, I. Clin. Exp. Allergy 2007, 37, 780–787. [Google Scholar] [CrossRef]
- Borna, E.; Nwaru, B.I.; Bjerg, A.; Mincheva, R.; Rådinger, M.; Lundbäck, B.; Ekerljung, L. Changes in the Prevalence of Asthma and Respiratory Symptoms in Western Sweden between 2008 and 2016. Allergy Eur. J. Allergy Clin. Immunol. 2019, 74, 1703–1715. [Google Scholar] [CrossRef] [PubMed]
- Oskouei, Y.M.; Hosseini, R.F.; Ahanchian, H.; Jarahi, L.; Ariaee, N.; Azad, F.J. Report of Common Aeroallergens among Allergic Patients in Northeastern Iran. Iran. J. Otorhinolaryngol. 2017, 29, 89. [Google Scholar]
- Forkel, S.; Beutner, C.; Schröder, S.S.; Bader, O.; Gupta, S.; Fuchs, T.; Schön, M.P.; Geier, J.; Buhl, T. Sensitization against Fungi in Patients with Airway Allergies over 20 Years in Germany. Int. Arch. Allergy Immunol. 2021, 182, 515–523. [Google Scholar] [CrossRef] [PubMed]
- De Vouge, M.W.; Thaker, A.J.; Zhang, L.; Muradia, G.; Rode, H.; Vijay, H.M. Molecular Cloning of IgE-Binding Fragments of Alternaria Alternata Allergens. Int. Arch. Allergy Immunol. 1998, 116, 261–268. [Google Scholar] [CrossRef]
- Kustrzeba-Wójcicka, I.; Siwak, E.; Terlecki, G.; Wolańczyk-Mędrala, A.; Mędrala, W. Alternaria Alternata and Its Allergens: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2014, 43, 354–365. [Google Scholar] [CrossRef]
- Achatz, G.; Oberkofler, H.; Lechenauer, E.; Simon, B.; Unger, A.; Kandler, D.; Ebner, C.; Prillinger, H.; Kraft, D.; Breitenbach, M. Molecular Cloning of Major and Minor Allergens of Alternaria Alternata and Cladosporium Herbarum. Mol. Immunol. 1995, 32, 213–227. [Google Scholar] [CrossRef]
- Simon-Nobbe, B.; Probst, G.; Kajava, A.V.; Oberkofler, H.; Susani, M.; Crameri, R.; Ferreira, F.; Ebner, C.; Breitenbach, M. IgE-Binding Epitopes of Enolases, a Class of Highly Conserved Fungal Allergens. J. Allergy Clin. Immunol. 2000, 106, 887–895. [Google Scholar] [CrossRef]
- Schneider, P.B.; Denk, U.; Breitenbach, M.; Richter, K.; Schmid-Grendelmeier, P.; Nobbe, S.; Himly, M.; Mari, A.; Ebner, C.; Simon-Nobbe, B. Alternaria Alternata NADP*-Dependent Mannitol Dehydrogenase Is an Important Fungal Allergen. Clin. Exp. Allergy 2006, 36, 1513–1524. [Google Scholar] [CrossRef]
- Shankar, J.; Singh, B.P.; Gaur, S.N.; Arora, N. Recombinant Glutathione-S-Transferase a Major Allergen from Alternaria Alternata for Clinical Use in Allergy Patients. Mol. Immunol. 2006, 43, 1927–1932. [Google Scholar] [CrossRef]
- Weichel, M.; Schmid-Grendelmeier, P.; Flückiger, S.; Breitenbach, M.; Blaser, K.; Crameri, R. Nuclear Transport Factor 2 Represents a Novel Cross-Reactive Fungal Allergen. Allergy Eur. J. Allergy Clin. Immunol. 2003, 58, 198–206. [Google Scholar] [CrossRef]
- Rid, R.; Önder, K.; MacDonald, S.; Lang, R.; Hawranek, T.; Ebner, C.; Hemmer, W.; Richter, K.; Simon-Nobbe, B.; Breitenbach, M. Alternaria Alternata TCTP, a Novel Cross-Reactive Ascomycete Allergen. Mol. Immunol. 2009, 46, 3476–3487. [Google Scholar] [CrossRef]
- Portnoy, J.; Olson, I.; Pacheco, F.; Barnes, C. Affinity Purification of a Major Alternaria Allergen Using a Monoclonal Antibody. Ann. Allergy 1990, 65, 109–114. [Google Scholar] [PubMed]
- Tabar, A.I.; Prieto, L.; Alba, P.; Nieto, A.; Rodríguez, M.; Torrecillas, M.; Huertas, B.; Gómez, E.; Fernández, F.J.; Blanca, M.; et al. Double-Blind, Randomized, Placebo-Controlled Trial of Allergen-Specific Immunotherapy with the Major Allergen Alt a 1. J. Allergy Clin. Immunol. 2019, 144, 216–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, J.; Gupta, P.D.; Sridhara, S.; Singh, B.P.; Gaur, S.N.; Arora, N. Immunobiochemical Analysis of Cross-Reactive Glutathione-S-Transferase Allergen from Different Fungal Sources. Immunol. Investig. 2005, 34, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Arandia, M.; Silva-Navas, J.; Ramírez-Castillejo, C.; Cubells-Baeza, N.; Gómez-Casado, C.; Barber, D.; Pozo, J.C.; Melendi, P.G.; Pacios, L.F.; Diáz-Perales, A. Characterisation of a Flavonoid Ligand of the Fungal Protein Alt a 1. Sci. Rep. 2016. [Google Scholar] [CrossRef]
- Cramer, R.A.; Lawrence, C.B. Cloning of a Gene Encoding an Alt a 1 Isoallergen Differentially Expressed by the Necrotrophic Fungus Alternaria Brassicicola during Arabidopsis Infection. Appl. Environ. Microbiol. 2003, 69, 2361. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Mukherjee, P.K. Trichoderma Virens Alt a 1 Protein May Target Maize PR5/Thaumatin-like Protein to Suppress Plant Defence: An in Silico Analysis. Physiol. Mol. Plant Pathol. 2020, 112, 101551. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, Y.; Liang, Y.; Dong, Y.; Yang, X.; Qiu, D. Verticillium Dahliae PevD1, an Alt a 1-like Protein, Targets Cotton PR5-like Protein and Promotes Fungal Infection. J. Exp. Bot. 2019, 70, 613–626. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Casado, C.; Murua-García, A.; Garrido-Arandia, M.; González-Melendi, P.; Sánchez-Monge, R.; Barber, D.; Pacios, L.F.; Díaz-Perales, A. Alt a 1 from Alternaria Interacts with PR5 Thaumatin-like Proteins. FEBS Lett. 2014, 588, 1501–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Mukherjee, P.K. Trichoderma Virens Bys1 May Competitively Inhibit Its Own Effector Protein Alt a 1 to Stabilize the Symbiotic Relationship with Plant-Evidence from Docking and Simulation Studies. 3 Biotech 2021, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.; Paciência, I.; Farraia, M.; Cavaleiro Rufo, J.; Castro Mendes, F.; Delgado, L.; Viegas, C.; Moreira, A. Airborne Food and Aeroallergens Levels in Healthcare Settings. An Unaccounted but Potentially Relevant Source of Exposure? J. Investig. Allergol. Clin. Immunol. 2020, 31. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Arandia, M.; Bretones, J.; Gómez-Casado, C.; Cubells, N.; Díaz-Perales, A.; Pacios, L.F. Computational Study of PH-Dependent Oligomerization and Ligand Binding in Alt a 1, a Highly Allergenic Protein with a Unique Fold. J. Comput. Aided. Mol. Des. 2016, 30, 365–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chruszcz, M.; Chapman, M.D.; Osinski, T.; Solberg, R.; Demas, M.; Porebski, P.J.; Majorek, K.A.; Pomés, A.; Minor, W. Alternaria Alternata Allergen Alt a 1: A Unique β-Barrel Protein Dimer Found Exclusively in Fungi. J. Allergy Clin. Immunol. 2012, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurup, V.P.; Vijay, H.M.; Kumar, V.; Castillo, L.; Elms, N. IgE Binding Synthetic Peptides of Alt a 1, a Major Allergen of Alternaria Alternata. Peptides 2003, 24, 179–185. [Google Scholar] [CrossRef]
- Garrido-Arandia, M.; Gomez-Casado, C.; Díaz-Perales, A.; Pacios, L.F. Molecular Dynamics of Major Allergens from Alternaria, Birch Pollen and Peach. Mol. Inform. 2014, 33, 682–694. [Google Scholar] [CrossRef]
- Caraballo, L.; Valenta, R.; Puerta, L.; Pomés, A.; Zakzuk, J.; Fernandez-Caldas, E.; Acevedo, N.; Sanchez-Borges, M.; Ansotegui, I.; Zhang, L.; et al. The Allergenic Activity and Clinical Impact of Individual IgE-Antibody Binding Molecules from Indoor Allergen Sources. World Allergy Organ. J. 2020, 13. [Google Scholar] [CrossRef]
- Hellings, P.W.; Steelant, B. Epithelial Barriers in Allergy and Asthma. J. Allergy Clin. Immunol. 2020, 145, 1499–1509. [Google Scholar] [CrossRef]
- Wiesner, D.L.; Klein, B.S. Lung Epithelium: Barrier Immunity to Inhaled Fungi and Driver of Fungal-Associated Allergic Asthma. Curr. Opin. Microbiol. 2017, 40, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, S.; Ahmad, S.; Irshad, R.; Goyal, Y.; Rafat, S.; Siddiqui, N.; Dev, K.; Husain, M.; Ali, S.; Mohan, A.; et al. TLRs in Pulmonary Diseases. Life Sci. 2019, 233. [Google Scholar] [CrossRef] [PubMed]
- Hayes, T.; Rumore, A.; Howard, B.; He, X.; Luo, M.; Wuenschmann, S.; Chapman, M.; Kale, S.; Li, L.; Kita, H.; et al. Innate Immunity Induced by the Major Allergen Alt a 1 From the Fungus Alternaria Is Dependent Upon Toll-Like Receptors 2/4 in Human Lung Epithelial Cells. Front. Immunol. 2018, 1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartemes, K.R.; Kita, H. Innate and Adaptive Immune Responses to Fungi in the Airway. J. Allergy Clin. Immunol. 2018, 142, 353–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yee, M.C.; Nichols, H.L.; Polley, D.; Saifeddine, M.; Pal, K.; Lee, K.; Wilson, E.H.; Daines, M.O.; Hollenberg, M.D.; Boitano, S.; et al. Protease-Activated Receptor-2 Signaling through β-Arrestin-2 Mediates Alternaria Alkaline Serine Protease-Induced Airway Inflammation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L1042–L1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toki, S.; Goleniewska, K.; Zhang, J.; Zhou, W.; Newcomb, D.C.; Zhou, B.; Kita, H.; Boyd, K.L.; Peebles, R.S. TSLP and IL-33 Reciprocally Promote Each Other’s Lung Protein Expression and ILC2 Receptor Expression to Enhance Innate Type-2 Airway Inflammation. Allergy 2020, 75, 1606–1617. [Google Scholar] [CrossRef]
- Kim, H.-K.; Lund, S.; Baum, R.; Rosenthal, P.; Khorram, N.; Doherty, T.A. Innate Type 2 Response to Alternaria Extract Enhances Ryegrass-Induced Lung Inflammation. Int. Arch. Allergy Immunol. 2014, 163, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Matsuwaki, Y.; Wada, K.; White, T.; Moriyama, H.; Kita, H. Alternaria Fungus Induces the Production of GM-CSF, Interleukin-6 and Interleukin-8 and Calcium Signaling in Human Airway Epithelium through Protease-Activated Receptor 2. Int. Arch. Allergy Immunol. 2012, 158 Suppl 1, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Zhang, J.; Toki, S.; Goleniewska, K.; Norlander, A.E.; Newcomb, D.C.; Wu, P.; Boyd, K.L.; Kita, H.; Peebles, R.S. COX Inhibition Increases Alternaria -Induced Pulmonary Group 2 Innate Lymphoid Cell Responses and IL-33 Release in Mice. J. Immunol. 2020, 205, 1157–1166. [Google Scholar] [CrossRef]
- Kato, T.; Kouzaki, H.; Matsumoto, K.; Hosoi, J.; Shimizu, T. The Effect of Calprotectin on TSLP and IL-25 Production from Airway Epithelial Cells. Allergol. Int. 2017, 66, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Denis, O.; Vincent, M.; Havaux, X.; De Prins, S.; Treutens, G.; Huygen, K. Induction of the Specific Allergic Immune Response Is Independent of Proteases from the Fungus Alternaria Alternata. Eur. J. Immunol. 2013, 43, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Arandia, M.; Tome-Amat, J.; Pazos-Castro, D.; Esteban, V.; Escribese, M.M.; Hernández-Ramírez, G.; Yuste-Montalvo, A.; Barber, D.; Pacios, L.F.; Díaz-Perales, A. Interaction of Alt a 1 with SLC22A17 in the Airway Mucosa. Allergy Eur. J. Allergy Clin. Immunol. 2019, 74, 2167–2180. [Google Scholar] [CrossRef] [PubMed]
- Hartl, D.; Tirouvanziam, R.; Laval, J.; Greene, C.M.; Habiel, D.; Sharma, L.; Yildirim, A.Ö.; Dela Cruz, C.S.; Hogaboam, C.M. Innate Immunity of the Lung: From Basic Mechanisms to Translational Medicine. J. Innate Immun. 2018, 10, 487–501. [Google Scholar] [CrossRef]
- Joshi, N.; Walter, J.M.; Misharin, A.V. Alveolar Macrophages. Cell. Immunol. 2018, 330, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Draijer, C.; Peters-Golden, M. Alveolar Macrophages in Allergic Asthma: The Forgotten Cell Awakes. Curr. Allergy Asthma Rep. 2017, 17, 12. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, S.A.; Ku, J.Y.; Cho, W.K.; Shin, C.H. Comparison of Allergens and Symptoms in Patients with Allergic Rhinitis between 1990s and 2010s. Allergy Asthma Clin. Immunol. 2020, 16, 1–7. [Google Scholar] [CrossRef]
- Valero, A.; Quirce, S.; Dávila, I.; Delgado, J.; Domínguez-Ortega, J. Allergic Respiratory Disease: Different Allergens, Different Symptoms. Allergy 2017, 72, 1306–1316. [Google Scholar] [CrossRef]
- Shin, S.H.; Ye, M.K.; Lee, D.W.; Chae, M.H.; Choi, S.Y. Development and Immunopathological Characteristics of an Alternaria-Induced Chronic Rhinosinusitis Mouse Model. PLoS ONE 2020, 15. [Google Scholar] [CrossRef]
- Cavagnero, K.J.; Badrani, J.H.; Naji, L.H.; Amadeo, M.B.; Shah, V.S.; Gasparian, S.; Pham, A.; Wang, A.W.; Seumois, G.; Croft, M.; et al. Unconventional ST2- and CD127-Negative Lung ILC2 Populations Are Induced by the Fungal Allergen Alternaria Alternata. J. Allergy Clin. Immunol. 2019, 144, 1432–1435. [Google Scholar] [CrossRef] [Green Version]
- Loghmani, A.; Raoofi, R.; Ownagh, A.; Delirezh, N. Alternaria Alternata Acts on Human Monocyte-Derived Dendritic Cells to Mediate Th2/Th17 Polarisation. Allergol. Immunopathol. 2017, 45, 175–182. [Google Scholar] [CrossRef]
- Wada, K.; Kobayashi, T.; Matsuwaki, Y.; Moriyama, H.; Kita, H. Alternaria Inhibits Double-Stranded RNA-Induced Cytokine Production through Toll-Like Receptor 3. Int. Arch. Allergy Immunol. 2013, 161, 75–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Didehdar, M.; Khoshbayan, A.; Vesal, S.; Darban-Sarokhalil, D.; Razavi, S.; Chegini, Z.; Shariati, A. An Overview of Possible Pathogenesis Mechanisms of Alternaria Alternata in Chronic Rhinosinusitis and Nasal Polyposis. Microb. Pathog. 2021, 155. [Google Scholar] [CrossRef] [PubMed]
- Mathä, L.; Martinez-Gonzalez, I.; Steer, C.A.; Takei, F. The Fate of Activated Group 2 Innate Lymphoid Cells. Front. Immunol. 2021, 12, 671966. [Google Scholar] [CrossRef]
- Stier, M.T.; Zhang, J.; Goleniewska, K.; Cephus, J.Y.; Rusznak, M.; Wu, L.; Van Kaer, L.; Zhou, B.; Newcomb, D.C.; Peebles, R.S. IL-33 Promotes the Egress of Group 2 Innate Lymphoid Cells from the Bone Marrow. J. Exp. Med. 2018, 215, 263–281. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, C.M.; Snelgrove, R.J. Type 2 Immunity: Expanding Our View. Sci. Immunol. 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Bartemes, K.; Chen, C.-C.; Iijima, K.; Drake, L.; Kita, H. IL-33–Responsive Group 2 Innate Lymphoid Cells Are Regulated by Female Sex Hormones in the Uterus. J. Immunol. 2018, 200, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Bartemes, K.R.; Iijima, K.; Kobayashi, T.; Kephart, G.M.; McKenzie, A.N.; Kita, H. IL-33–Responsive Lineage − CD25 + CD44 Hi Lymphoid Cells Mediate Innate Type 2 Immunity and Allergic Inflammation in the Lungs. J. Immunol. 2012, 188, 1503–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valladao, A.C.; Frevert, C.W.; Koch, L.K.; Campbell, D.J.; Ziegler, S.F. STAT6 Regulates the Development of Eosinophilic versus Neutrophilic Asthma in Response to Alternaria Alternata. J. Immunol. 2016, 197, 4541–4551. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, A.K.; Makhija, S.; Stahr, N.; Sandey, M.; Suryawanshi, A.; Mishra, A. Pyruvate Kinase M2 in Lung APCs Regulates Alternaria-Induced Airway Inflammation. Immunobiology 2020, 225, 151956. [Google Scholar] [CrossRef]
- Bousquet, J.; Pfaar, O.; Agache, I.; Bedbrook, A.; Akdis, C.A.; Canonica, G.W.; Chivato, T.; Al-Ahmad, M.; Latiff, A.H.A.; Ansotegui, I.J.; et al. ARIA-EAACI Care Pathways for Allergen Immunotherapy in Respiratory Allergy. Clin. Transl. Allergy 2021, 11, e12014. [Google Scholar] [CrossRef]
- Lombardi, C.; Savi, E.; Ridolo, E.; Passalacqua, G.; Canonica, G.W. Is Allergic Sensitization Relevant in Severe Asthma? Which Allergens May Be Culprits? World Allergy Organ. J. 2017, 10, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vélez-Pereira, A.M.; De Linares, C.; Canela, M.-A.; Belmonte, J. Logistic Regression Models for Predicting Daily Airborne Alternaria and Cladosporium Concentration Levels in Catalonia (NE Spain). Int. J. Biometeorol. 2019, 63, 1541–1553. [Google Scholar] [CrossRef] [PubMed]
- Feo Brito, F.; Alonso, A.M.; Carnés, J.; Martín-Martín, R.; Fernández-Caldas, E.; Galindo, P.A.; Alfaya, T.; Amo-Salas, M. Alternaria Alternata Spores, Alt a 1, and Symptoms Correlation Between Alt a 1 Levels and Clinical Symptoms in Alternaria Alternata-Monosensitized Patients. J. Investig. Allergol. Clin. Immunol. 2012, 22, 154–159. [Google Scholar]
- Hernández-Ramírez, G.; Pazos-Castro, D.; Gómez Torrijos, E.; Yuste Montalvo, A.; Romero-Sahagun, A.; González-Klein, Z.; Jimeno-Nogales, L.; Escribese, M.M.; Extremera Ortega, A.; Nuñez-Borque, E.; et al. Group 1 Allergens, Transported by Mold Spores, Induce Asthma Exacerbation in a Mouse Model. Allergy Eur. J. Allergy Clin. Immunol. 2020, 75, 2388–2391. [Google Scholar] [CrossRef]
- Nasser, S.M.; Pulimood, T.B. Allergens and Thunderstorm Asthma. Curr. Allergy Asthma Rep. 2009, 9, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Gorska, M.M. Mouse Models of Asthma. Methods Mol. Biol. 2018, 1809, 351–362. [Google Scholar] [CrossRef]
- Kobayashi, T.; Iijima, K.; Radhakrishnan, S.; Mehta, V.; Vassallo, R.; Lawrence, C.B.; Cyong, J.-C.; Pease, L.R.; Oguchi, K.; Kita, H. Asthma-Related Environmental Fungus, Alternaria, Activates Dendritic Cells and Produces Potent Th2 Adjuvant Activity. J. Immunol. 2009, 182, 2502–2510. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, M.F.; Uriel, N.; Teifoori, F.; Postigo, I.; Suñén, E.; Martínez, J. The Major Alternaria Alternata Allergen, Alt a 1: A Reliable and Specific Marker of Fungal Contamination in Citrus Fruits. Int. J. Food Microbiol. 2017, 257, 26–30. [Google Scholar] [CrossRef]
- Popescu, F.-D. Cross-Reactivity between Aeroallergens and Food Allergens. World J. Methodol. 2015, 5, 31. [Google Scholar] [CrossRef]
- Herrera, I.; Moneo, I.; Caballero, M.L.; De Paz, S.; Perez Pimiento, A.; Rebollo, S. Food Allergy to Spinach and Mushroom. Allergy Eur. J. Allergy Clin. Immunol. 2002, 57, 261–262. [Google Scholar] [CrossRef]
- Di Bona, D.; Frisenda, F.; Albanesi, M.; Di Lorenzo, G.; Caiaffa, M.F.; Macchia, L. Efficacy and Safety of Allergen Immunotherapy in Patients with Allergy to Molds: A Systematic Review. Clin. Exp. Allergy 2018, 48, 1391–1401. [Google Scholar] [CrossRef]
- Jutel, M.; Agache, I.; Bonini, S.; Burks, A.W.; Calderon, M.; Canonica, W.; Cox, L.; Demoly, P.; Frew, A.J.; O’Hehir, R.; et al. International Consensus on Allergy Immunotherapy. J. Allergy Clin. Immunol. 2015, 136, 556–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderon, M.A.; Demoly, P.; Casale, T.; Akdis, C.A.; Bachert, C.; Bewick, M.; Bilò, B.M.; Bohle, B.; Bonini, S.; Bush, A.; et al. Allergy Immunotherapy across the Life Cycle to Promote Active and Healthy Ageing: From Research to Policies: An AIRWAYS Integrated Care Pathways (ICPs) Programme Item (Action Plan B3 of the European Innovation Partnership on Active and Healthy Ageing) An. Clin. Transl. Allergy 2016, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, S.; Sprünken, A.; Wagner, N.; Tenbrock, K.; Ott, H. Clinical Relevance of IgE-Mediated Sensitization against the Mould Alternaria Alternata in Children with Asthma. Ther. Adv. Respir. Dis. 2017, 11, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soffer, N.; Green, B.J.; Acosta, L.; Divjan, A.; Sobek, E.; Lemons, A.R.; Rundle, A.G.; Jacobson, J.S.; Goldstein, I.F.; Miller, R.L.; et al. Alternaria Is Associated with Asthma Symptoms and Exhaled NO among NYC Children. J. Allergy Clin. Immunol. 2018, 142, 1366–1368. [Google Scholar] [CrossRef] [PubMed]
- Pfaar, O.; Agache, I.; de Blay, F.; Bonini, S.; Chaker, A.M.; Durham, S.R.; Gawlik, R.; Hellings, P.W.; Jutel, M.; Kleine-Tebbe, J.; et al. Perspectives in Allergen Immunotherapy: 2019 and Beyond. Allergy 2019, 74, 3–25. [Google Scholar] [CrossRef]
- Gada, S.; Haymore, B.; McCoy, L.; Kosisky, S.; Nelson, M. Frequency of Mold and Pollen Mixing in Allergen Immunotherapy Prescriptions within a Large Health Care System, 1990-2010. J. Allergy Clin. Immunol. 2012, 129, 1151–1153. [Google Scholar] [CrossRef]
- Zapatero, L.; Martínez-Cañavate, A.; Lucas, J.M.; Guallar, I.; Torres, J.; Guardia, P.; de La Torre, F.; Pedemonte, C. Clinical Evolution of Patients with Respiratory Allergic Disease Due to Sensitisation to Alternaria Alternata Being Treated with Subcutaneous Immunotherapy. Allergol. Immunopathol. 2011, 39, 79–84. [Google Scholar] [CrossRef]
- Martínez-Cañavate Burgos, A.; Valenzuela-Soria, A.; Rojo-Hernández, A. Immunotherapy with Alternaria Alternata: Present and Future. Allergol. Immunopathol. 2007, 35, 259–263. [Google Scholar] [CrossRef]
- Sanchez, H.; Bush, R.K. A Review of Alternaria Alternata Sensitivity. Rev. Iberoam. Micol. 2001, 18, 56–59. [Google Scholar]
- Kiliç, M.; Altintaş, D.U.; Yilmaz, M.; Bingöl-Karakoç, G.; Burgut, R.; Güneşer-Kendirli, S. Evaluation of Efficacy of Immunotherapy in Children with Asthma Monosensitized to Alternaria. Turk. J. Pediatr. 2011, 53, 285–294. [Google Scholar] [PubMed]
- Aden, E.; Weber, B.; Bossert, J.; Teppke, M.; Frank, E.; Wahl, R.; Fiebig, H.; Cromwell, O. Standardization of Alternaria Alternata: Extraction and Quantification of Alt a 1 by Using an MAb-Based 2-Site Binding Assay. J. Allergy Clin. Immunol. 1999, 104, 128–135. [Google Scholar] [CrossRef]
- Prieto, L.; Palacios, R.; Aldana, D.; Ferrer, A.; Perez-Frances, C.; Lopez, V.; Rojas, R. Effect of Allergen-Specific Immunotherapy with Purified Alt A1 on AMP Responsiveness, Exhaled Nitric Oxide and Exhaled Breath Condensate PH: A Randomized Double Blind Study. Allergy Asthma Clin. Immunol. Off. J. Can. Soc. Allergy Clin. Immunol. 2010, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Lizaso, M.T.; Martínez, A.; Asturias, J.A.; Algorta, J.; Madariaga, B.; Labarta, N.; Tabar, A.I. Biological Standardization and Maximum Tolerated Dose Estimation of an Alternaria Alternata Allergenic Extract. J. Investig. Allergol. Clin. Immunol. 2006, 16, 94–103. [Google Scholar] [PubMed]
- Lizaso, M.T.; Tabar, A.I.; García, B.E.; Gómez, B.; Algorta, J.; Asturias, J.A.; Martínez, A. Double-Blind, Placebo-Controlled Alternaria Alternata Immunotherapy: In Vivo and in Vitro Parameters. Pediatr. Allergy Immunol. Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2008, 19, 76–81. [Google Scholar] [CrossRef]
- Serna-Candel, C.; Moreno-Perez, O.; Soriano, V.; Martínez, A. Churg-Strauss Syndrome Triggered by Hyposensitization to Alternaria Fungus. Clin. Rheumatol. 2007, 26, 2195–2196. [Google Scholar] [CrossRef]
- Tabar, A.I.; Lizaso, M.T.; García, B.E.; Gómez, B.; Echechipía, S.; Aldunate, M.T.; Madariaga, B.; Martínez, A. Double-Blind, Placebo-Controlled Study of Alternaria Alternata Immunotherapy: Clinical Efficacy and Safety. Pediatr. Allergy Immunol. 2008, 19, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Cañavate, A.; Eseverri, J.L.; Ródenas, R.; Tabar, A.I.; Gardee, J.; Torres, J.; Boné, J.; Pedemonte, C. Evaluation of Paediatric Tolerance to an Extract of Alternaria Alternata under Two Treatment Regimes. A Multicentre Study. Allergol. Immunopathol. 2005, 33, 138–141. [Google Scholar] [CrossRef]
Figure 1.
Protein structure of Alt a 1: (a) Schematic diagram of the dimeric structure of Alt a 1 in its “butterfly” shape; (b) molecular surface of the dimeric structure of Alt a 1. Chains are shown in different colors and the position of the epitopes are shown in yellow, the epitopes are oriented to the outside of the structure leading the interaction with IgE antibodies; (c) schematic diagram of the tetrameric structure of Alt a 1; (d) molecular surface of the tetrameric structure of Alt a 1, chains are shown in different colors and the epitopes in yellow; (e) detail of the tetrameric molecular surface, showing the IgE binding epitopes (yellow) buried in the structure. PDB code, 3v0r.
Figure 1.
Protein structure of Alt a 1: (a) Schematic diagram of the dimeric structure of Alt a 1 in its “butterfly” shape; (b) molecular surface of the dimeric structure of Alt a 1. Chains are shown in different colors and the position of the epitopes are shown in yellow, the epitopes are oriented to the outside of the structure leading the interaction with IgE antibodies; (c) schematic diagram of the tetrameric structure of Alt a 1; (d) molecular surface of the tetrameric structure of Alt a 1, chains are shown in different colors and the epitopes in yellow; (e) detail of the tetrameric molecular surface, showing the IgE binding epitopes (yellow) buried in the structure. PDB code, 3v0r.

Figure 2.
Immune responses induced in airway epithelium by Alternaria alternata. Spores and mycelium fragments containing allergenic compounds (such as Alt a 1) can reach upper airways, where they can interact with TLRs (TLR2, TLR4), PAR 2, or SLC22A17 receptors present in the alveolar macrophages and epithelial cells. The activation of these types of cells induces proinflammatory responses by means of the production of alarmins (IL-33, IL-25, TSLP) and other proinflammatory cytokines. Alarmins promote the recruitment and activation of innate lymphoid cells type 2 (ILC2), which have an important role in the development of Alternaria-induced type 2 responses (e.g., eosinophil infiltration, mucus hypersecretion, airway hyperreactivity, IgE production, etc.). EOS, eosinophil; NEU, neutrophil; DC, dendritic cell; AM, alveolar macrophage.
Figure 2.
Immune responses induced in airway epithelium by Alternaria alternata. Spores and mycelium fragments containing allergenic compounds (such as Alt a 1) can reach upper airways, where they can interact with TLRs (TLR2, TLR4), PAR 2, or SLC22A17 receptors present in the alveolar macrophages and epithelial cells. The activation of these types of cells induces proinflammatory responses by means of the production of alarmins (IL-33, IL-25, TSLP) and other proinflammatory cytokines. Alarmins promote the recruitment and activation of innate lymphoid cells type 2 (ILC2), which have an important role in the development of Alternaria-induced type 2 responses (e.g., eosinophil infiltration, mucus hypersecretion, airway hyperreactivity, IgE production, etc.). EOS, eosinophil; NEU, neutrophil; DC, dendritic cell; AM, alveolar macrophage.

Figure 3.
Co-sensitization phenomena between Alternaria and other allergens: (a) asthma exacerbation by sensitization to Alternaria grass pollen. In Central Spain, grass pollen patients suffer from asthma during late summer and early fall, when pollen is no longer present. In contrast, in this season, Alternaria spore peaks were registered (data from Ciudad Real, 2020, SEAIC, https://www.polenes.com/es/home, accessed on 1 October 2021). Considering that thunderstorms and grass postharvest are common at this time, the authors described the role of spores as Group-1 grass allergen carriers and suggested that the inhalation of loaded spores may be the cause of the asthma exacerbations in grass pollen patients; (b) Alternaria, kiwifruit co-sensitization. Alternaria infects kiwifruits secreting Alt a 1 that can be detected in pulp at 5 days post infection (dpi) while fungal hypha is not visible. The partial inhibition of a PR5 thaumatin-like protein (Act d 2) by Alt a 1 was reported, evidencing the role of the fungal protein in plant pathogenesis, and providing an explanation for the co-relation between Alt a 1 and Act d 2 sensitizations as a result of consumption of Alternaria-infected kiwifruits but apparently in good conditions.
Figure 3.
Co-sensitization phenomena between Alternaria and other allergens: (a) asthma exacerbation by sensitization to Alternaria grass pollen. In Central Spain, grass pollen patients suffer from asthma during late summer and early fall, when pollen is no longer present. In contrast, in this season, Alternaria spore peaks were registered (data from Ciudad Real, 2020, SEAIC, https://www.polenes.com/es/home, accessed on 1 October 2021). Considering that thunderstorms and grass postharvest are common at this time, the authors described the role of spores as Group-1 grass allergen carriers and suggested that the inhalation of loaded spores may be the cause of the asthma exacerbations in grass pollen patients; (b) Alternaria, kiwifruit co-sensitization. Alternaria infects kiwifruits secreting Alt a 1 that can be detected in pulp at 5 days post infection (dpi) while fungal hypha is not visible. The partial inhibition of a PR5 thaumatin-like protein (Act d 2) by Alt a 1 was reported, evidencing the role of the fungal protein in plant pathogenesis, and providing an explanation for the co-relation between Alt a 1 and Act d 2 sensitizations as a result of consumption of Alternaria-infected kiwifruits but apparently in good conditions.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hernandez-Ramirez, G.; Barber, D.; Tome-Amat, J.; Garrido-Arandia, M.; Diaz-Perales, A. Alternaria as an Inducer of Allergic Sensitization. J. Fungi 2021, 7, 838. https://doi.org/10.3390/jof7100838
AMA Style
Hernandez-Ramirez G, Barber D, Tome-Amat J, Garrido-Arandia M, Diaz-Perales A. Alternaria as an Inducer of Allergic Sensitization. Journal of Fungi. 2021; 7(10):838. https://doi.org/10.3390/jof7100838
Chicago/Turabian StyleHernandez-Ramirez, Guadalupe, Domingo Barber, Jaime Tome-Amat, Maria Garrido-Arandia, and Araceli Diaz-Perales. 2021. "Alternaria as an Inducer of Allergic Sensitization" Journal of Fungi 7, no. 10: 838. https://doi.org/10.3390/jof7100838
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.