Molecular Basis and Clinical Application of Growth-Factor-Independent In Vitro Myeloid Colony Formation in Chronic Myelomonocytic Leukemia

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
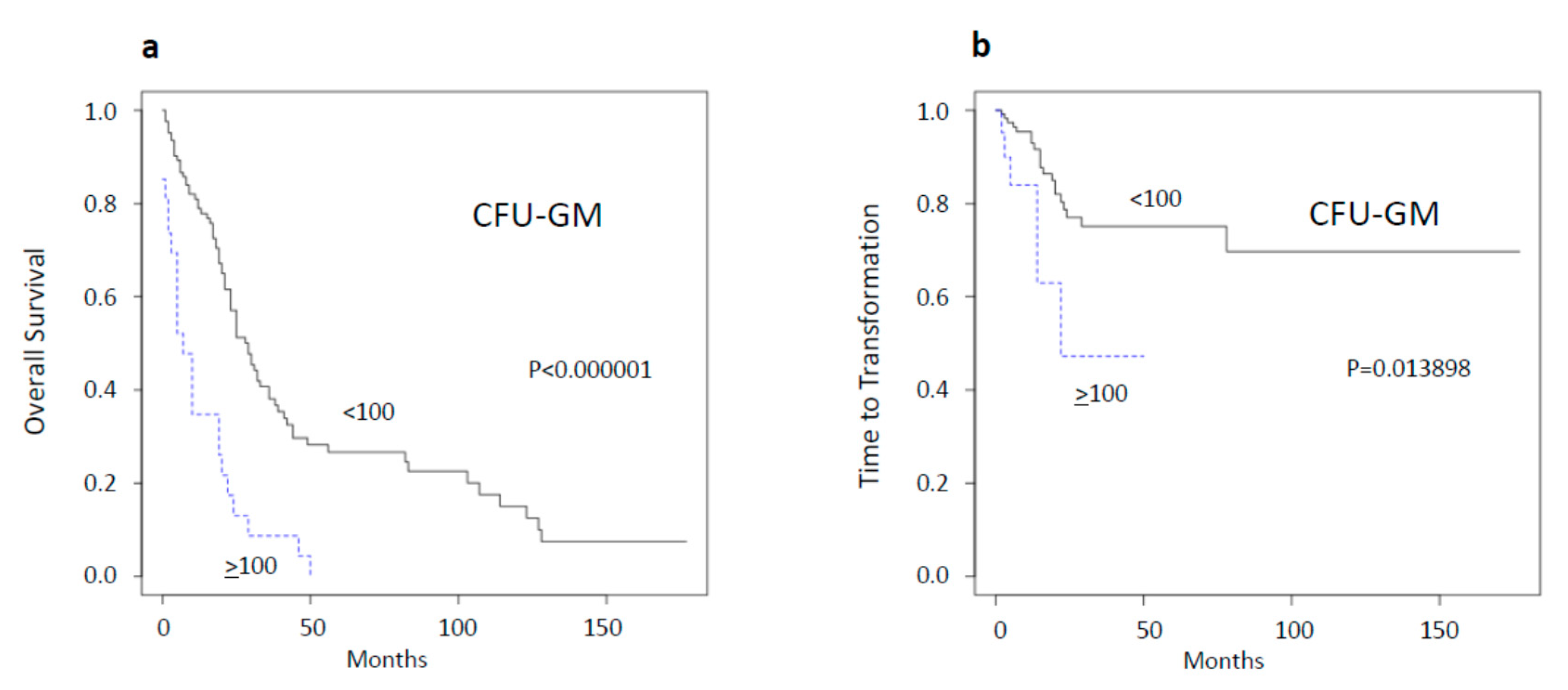
2.1. Spontaneous Myeloid Colony Growth Is a Strong Predictor of Survival and Time to Transformation
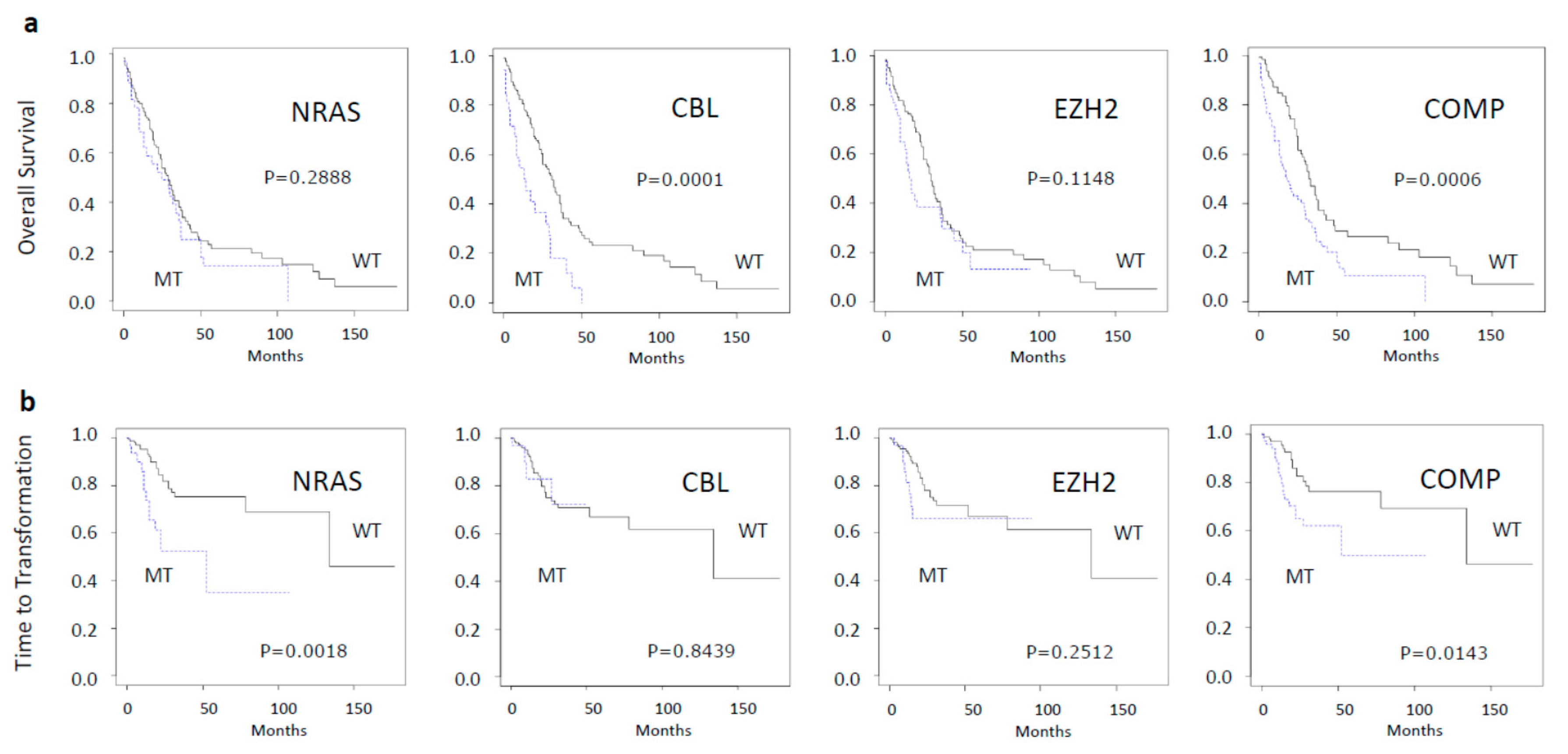
2.2. Impact of Molecular Aberrations on Spontaneous Myeloid Colony Growth
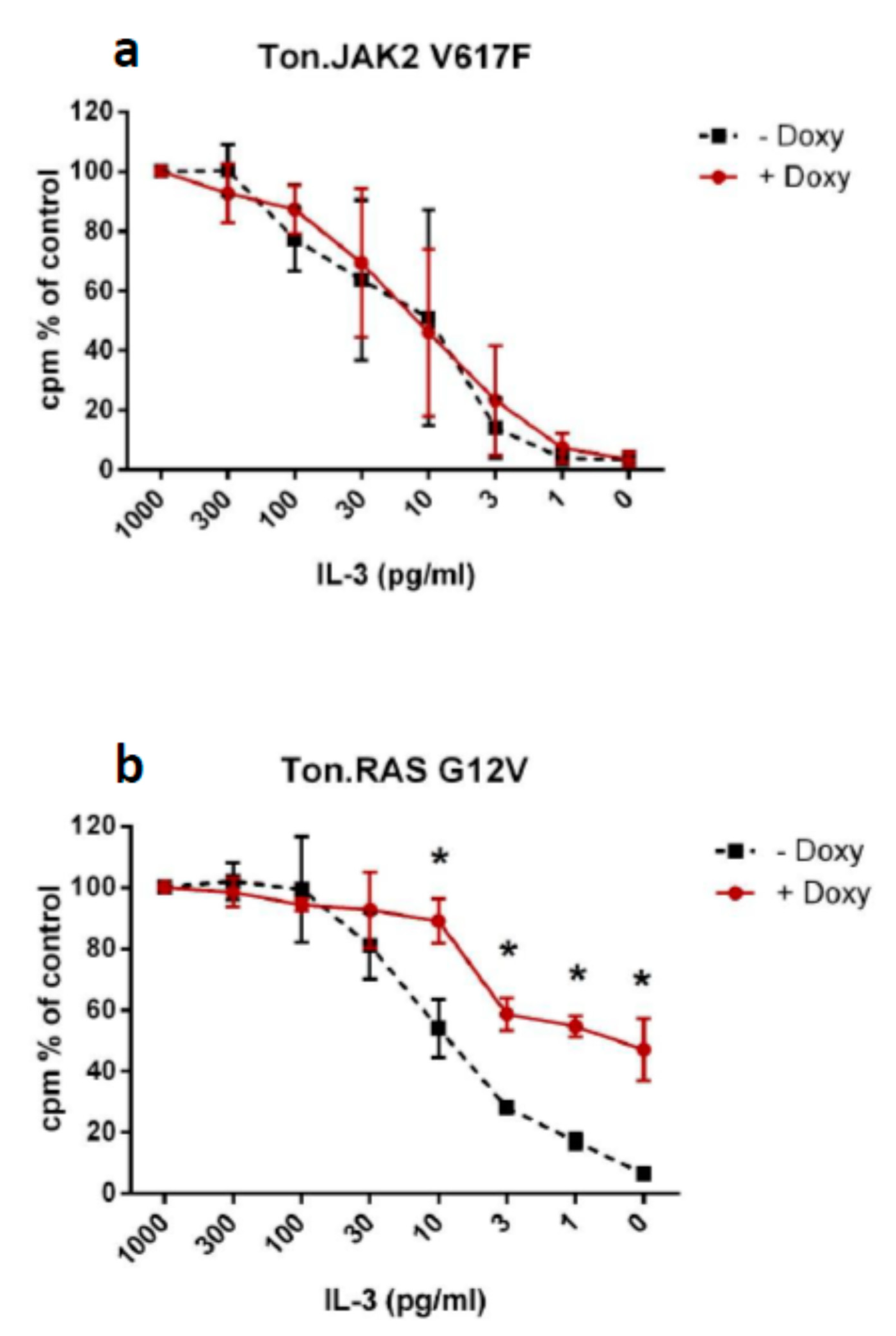
2.3. Impact of RAS G12V and JAK2 V617F on Growth Factor Dependence in Ba/F3 Cells
2.4. Generation of an in Vitro Myeloid Colony Growth Derived Molecular Pattern and Its Clinical Implication
3. Discussion
4. Patients and Methods
4.1. Patients
4.2. Colony Assay
4.3. Molecular Studies
4.4. Ba/F3 Cells with Inducible Expression of RAS G12V and JAK2 V617F
4.5. Measurement of 3H-thymidine Uptake
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Lowenberg, B.; van Putten, W.; Touw, I.P.; Delwel, R.; Santini, V. Autonomous Proliferation of Leukemic Cells in Vitro as a Determinant of Prognosis in Adult Acute Myeloid Leukemia. N. Engl. J. Med. 1993, 328, 614–619. [Google Scholar] [CrossRef]
- Dittrich, C.; Dittrich, E.; Sevelda, P.; Hudec, M.; Salzer, H.; Grunt, T.; Eliason, J. Clonogenic Growth in Vitro: An Independent Biologic Prognostic Factor in Ovarian Carcinoma. J. Clin. Oncol. 1991, 9, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Geissler, K.; Hinterberger, W.; Bettelheim, P.; Haas, O.; Lechner, K. Colony Growth Characteristics in Chronic Myelomonocytic Leukemia. Leuk. Res. 1988, 12, 373–377. [Google Scholar] [CrossRef]
- Everson, M.P.; Brown, C.B.; Lilly, M.B. Interleukin-6 and Granulocyte-Macrophage Colony-Stimulating Factor Are Candidate Growth Factors for Chronic Myelomonocytic Leukemia Cells. Blood 1989, 74, 1472–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramshaw, H.S.; Bardy, P.G.; Lee, M.A.; Lopez, A.F. Chronic Myelomonocytic Leukemia Requires Granulocyte-Macrophage Colony-Stimulating Factor for Growth in Vitro and in Vivo. Exp. Hematol. 2002, 30, 1124–1131. [Google Scholar] [CrossRef]
- Sagaster, V.; Öhler, L.; Berer, A.; Kabrna, E.; Ofner, P.; Lechner, K.; Geissler, K. High Spontaneous Colony Growth in Chronic Myelomonocytic Leukemia Correlates with Increased Disease Activity and Is a Novel Prognostic Factor for Predicting Short Survival. Ann. Hematol. 2004, 83, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Geissler, K.; Jäger, E.; Barna, A.; Alendar, T.; Ljubuncic, E.; Sliwa, T.; Valent, P. Chronic Myelomonocytic Leukemia Patients with RAS Pathway Mutations Show High in Vitro Myeloid Colony Formation in the Absence of Exogenous Growth Factors. Leukemia 2016, 30, 2280–2281. [Google Scholar] [CrossRef] [PubMed]
- Geissler, K.; Jäger, E.; Barna, A.; Gurbisz, M.; Graf, T.; Graf, E.; Nösslinger, T.; Pfeilstöcker, M.; Tüchler, H.; Sliwa, T.; et al. Correlation of RAS-Pathway Mutations and Spontaneous Myeloid Colony Growth with Progression and Transformation in Chronic Myelomonocytic Leukemia—A Retrospective Analysis in 337 Patients. Int. J. Mol. Sci. 2020, 21, 3025. [Google Scholar] [CrossRef]
- Bradley, T.; Metcalf, D. The growth of mouse bone marrow cells in vitro. Aust. J. Exp. Biol. Med. 1966, 44, 287–300. [Google Scholar] [CrossRef]
- Metcalf, D. Regulation of Hemopoiesis. Nouv. Rev. Fr. Hematol. 1978, 20, 521–533. [Google Scholar] [PubMed]
- Öhler, L.; Geissler, K.; Hinterberger, W. Diagnostic and Prognostic Value of Colony Formation of Hematopoietic Progenitor Cells in Myeloid Malignancies. Wien Klein. Wochenschr. 2003, 115, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.; Li, Z.; Du, J.; Ryu, M.-J.; Taylor, P.R.; Fleming, M.D.; Young, K.H.; Pitot, H.; Zhang, J. Endogenous Oncogenic Nras Mutation Promotes Aberrant GM-CSF Signaling in Granulocytic/Monocytic Precursors in a Murine Model of Chronic Myelomonocytic Leukemia. Blood 2010, 116, 5991–6002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Meter, M.E.M.; Díaz-Flores, E.; Archard, J.A.; Passegué, E.; Irish, J.M.; Kotecha, N.; Nolan, G.P.; Shannon, K.; Braun, B.S. K-RasG12D Expression Induces Hyperproliferation and Aberrant Signaling in Primary Hematopoietic Stem/Progenitor Cells. Blood 2007, 109, 3945–3952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Haigis, K.M.; McDaniel, A.; Harding-Theobald, E.; Kogan, S.C.; Akagi, K.; Wong, J.C.Y.; Braun, B.S.; Wolff, L.; Jacks, T.; et al. Hematopoiesis and Leukemogenesis in Mice Expressing Oncogenic NrasG12D from the Endogenous Locus. Blood 2011, 117, 2022–2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, R.J.; Leedy, M.B.; Munugalavadla, V.; Voorhorst, C.S.; Li, Y.; Yu, M.; Kapur, R. Human Somatic PTPN11 Mutations Induce Hematopoietic-Cell Hypersensitivity to Granulocyte-Macrophage Colony-Stimulating Factor. Blood 2005, 105, 3737–3742. [Google Scholar] [CrossRef] [Green Version]
- Le, D.T.; Kong, N.; Zhu, Y.; Lauchle, J.O.; Aiyigari, A.; Braun, B.S.; Wang, E.; Kogan, S.C.; Le Beau, M.M.; Parada, L.; et al. Somatic Inactivation of Nf1 in Hematopoietic Cells Results in a Progressive Myeloproliferative Disorder. Blood 2004, 103, 4243–4250. [Google Scholar] [CrossRef] [Green Version]
- Fedi, P.; Tronick, S.R.; Aaronson, S.A. Growth Factors. In Cancer Medicine; Holland, J.F., Bast, R.C., Morton, D.L., Frei, E., Kufe, D.W., Weichselbaum, R.R., Eds.; Williams and Wilkins: Baltimore, MD, USA, 1997; pp. 41–64. [Google Scholar]
- Geissler, K.; Ohler, L.; Födinger, M.; Virgolini, I.; Leimer, M.; Kabrna, E.; Kollars, M.; Skoupy, S.; Bohle, B.; Rogy, M.; et al. Interleukin 10 Inhibits Growth and Granulocyte/Macrophage Colony-Stimulating Factor Production in Chronic Myelomonocytic Leukemia Cells. J. Exp. Med. 1996, 184, 1377–1384. [Google Scholar] [CrossRef]
- Geissler, K.; Jäger, E.; Barna, A.; Gurbisz, M.; Marschon, R.; Graf, T.; Graf, E.; Borjan, B.; Jilch, R.; Geissler, C.; et al. The Austrian Biodatabase for Chronic Myelomonocytic Leukemia (ABCMML): A Representative and Useful Real-Life Data Source for Further Biomedical Research. Wien Klein. Wochenschr. 2019, 131, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 Revision to the World Health Organization Classification of Myeloid Neoplasms and Acute Leukemia. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef]
- Li, M.M.; Datto, M.; Duncavage, E.J.; Kulkarni, S.; Lindeman, N.I.; Roy, S.; Tsimberidou, A.M.; Vnencak-Jones, C.L.; Wolff, D.J.; Younes, A.; et al. Standards and Guidelines for the Interpretation and Reporting of Sequence Variants in Cancer. J. Mol. Diagn. 2017, 19, 4–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoermann, G.; Blatt, K.; Greiner, G.; Putz, E.M.; Berger, A.; Herrmann, H.; Cerny-Reiterer, S.; Gleixner, K.V.; Walz, C.; Hoetzenecker, K.; et al. CD52 Is a Molecular Target in Advanced Systemic Mastocytosis. FASEB J. 2014, 28, 3540–3551. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Factors | Factors Present Md OS (mo) | Factors Absent Md OS (mo) | Chi-Square Statistics | p-Value |
|---|---|---|---|---|
| CFU-GM ≥ 100/105 MNC | 7 | 29 | 28.082379 | 1.162596 × 10−7 |
| WBC ≥ 13 G/L | 20 | 29 | 9.922073 | 0.001633 |
| Hb < 10 g/dL | 17 | 25 | 7.288609 | 0.006939 |
| PLT < 100 G/L | 19 | 29 | 9.656587 | 0.001887 |
| PB Blasts present | 17 | 29 | 18.088790 | 0.000021 |
| - | Med LFS | Med LFS | - | - |
| CFU-GM ≥ 100/105 MNC | 22 | - | 6.050997 | 0.013898 |
| WBC ≥ 13 G/L | - | - | 0.520107 | 0.470796 |
| Hb < 10 g/dL | - | - | 0.609919 | 0.434818 |
| PLT < 100 G/L | - | - | 0.320680 | 0.571199 |
| PB Blasts present | - | - | 6.702843 | 0.009626 |
| Parameter | Hazard Ratio | 95% Confidence Interval | p-Value |
|---|---|---|---|
| CFU-GM ≥ 100/105 MNC | 1.485 | 1.145–1.1926 | 0.003 |
| WBC ≥ 13 G/L | 1.515 | 0.946–2.426 | 0.084 |
| Hb < 10 g/dL | 1.348 | 0.818–2.222 | 0.241 |
| PLT < 100 G/L | 1.602 | 1.060–2.422 | 0.025 |
| PB Blasts present | 1.413 | 0.850–2.351 | 0.183 |
| Genes | Mutated Samples Md CFU-GM (Q1–Q3) | Wildtype Samples Md CFU-GM (Q1–Q3) | p-Value Mann-Whitney |
|---|---|---|---|
| NRAS | 176 (25–264) | 7 (0–32) | <00001 |
| CBL | 35 (17–229) | 10 (0–63) | 0.0114 |
| EZH2 | 39 (8–389) | 10 (1–58) | 0.02926 |
| ASXL1 | 31 (2–204) | 9 (1–51) | 0.05118 |
| RUNX1 | 113 (2–247) | 10 (1–52) | 0.05876 |
| SF3B1 | 194 (0–285) | 11 (1–63) | 0.29834 |
| KRAS | 17 (1–76) | 11 (1–109) | 0.75656 |
| SRSF2 | 8 (1–136) | 12 (1–84) | 0.86502 |
| NF1 | 4 (0–216) | 11 (1–99) | 0.71884 |
| DNMT3A | 6 (0–139) | 13 (1–104) | 0.57548 |
| ZRSR2 | 15 (0–40) | 12 (1–109) | 0.47152 |
| SETBP1 | 6 (0–173) | 13 (1–98) | 0.42952 |
| TP53 | 4 (0–61) | 12 (1–109) | 0.33706 |
| U2AF1 | 9 (0–33) | 13 (1–109) | 0.29834 |
| PTPN11 | 0 (0–180) | 13 (1–102) | 0.11876 |
| IDH | 0 (0–12) | 13 (1–109) | 0.03156 |
| TET2 | 9 (0–52) | 37 (4–195) | 0.02382 |
| JAK2 | 3 (0–11) | 16 (1–116) | 0.00854 |
| Genes | Regression Coefficient B | S.D. | 2-Tail p-Value |
|---|---|---|---|
| NRAS | 4.51 | 1.06 | 4.57 × 10−5 |
| NF1 | 4.26 | 1.54 | 0.0067 |
| CBL | 2.27 | 0.95 | 0.0188 |
| EZH2 | 2.05 | 0.83 | 0.0154 |
| SF3B1 | 4.38 | 3.45 | 0.2073 |
| KRAS | 2.45 | 1.99 | 0.2212 |
| RUNX1 | 1.83 | 1.61 | 0.2580 |
| ASXL1 | 0.87 | 1.16 | 0.4546 |
| U2AF1 | 0.49 | 1.51 | 0.7441 |
| PTPN11 | 0.13 | 2.57 | 0.9584 |
| SETBP1 | 0.05 | 0.73 | 0.9408 |
| SRSF2 | −0.15 | 0.95 | 0.8748 |
| JAK2 | −0.85 | 0.89 | 0.3406 |
| IDH1/2 | −1.09 | 2.39 | 0.6495 |
| ZRSR2 | −1.03 | 0.90 | 0.2541 |
| TP53 | −1.82 | 2.95 | 0.5387 |
| DNMT3A | −1.97 | 1.50 | 0.1937 |
| TET2 | −1.29 | 0.64 | 0.0484 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geissler, K.; Jäger, E.; Barna, A.; Gurbisz, M.; Graf, T.; Graf, E.; Nösslinger, T.; Pfeilstöcker, M.; Machherndl-Spandl, S.; Stauder, R.; et al. Molecular Basis and Clinical Application of Growth-Factor-Independent In Vitro Myeloid Colony Formation in Chronic Myelomonocytic Leukemia. Int. J. Mol. Sci. 2020, 21, 6057. https://doi.org/10.3390/ijms21176057
Geissler K, Jäger E, Barna A, Gurbisz M, Graf T, Graf E, Nösslinger T, Pfeilstöcker M, Machherndl-Spandl S, Stauder R, et al. Molecular Basis and Clinical Application of Growth-Factor-Independent In Vitro Myeloid Colony Formation in Chronic Myelomonocytic Leukemia. International Journal of Molecular Sciences. 2020; 21(17):6057. https://doi.org/10.3390/ijms21176057
Chicago/Turabian StyleGeissler, Klaus, Eva Jäger, Agnes Barna, Michael Gurbisz, Temeida Graf, Elmir Graf, Thomas Nösslinger, Michael Pfeilstöcker, Sigrid Machherndl-Spandl, Reinhard Stauder, and et al. 2020. "Molecular Basis and Clinical Application of Growth-Factor-Independent In Vitro Myeloid Colony Formation in Chronic Myelomonocytic Leukemia" International Journal of Molecular Sciences 21, no. 17: 6057. https://doi.org/10.3390/ijms21176057