
Natural Occurring Silks and Their Analogues as Materials for Nerve Conduits

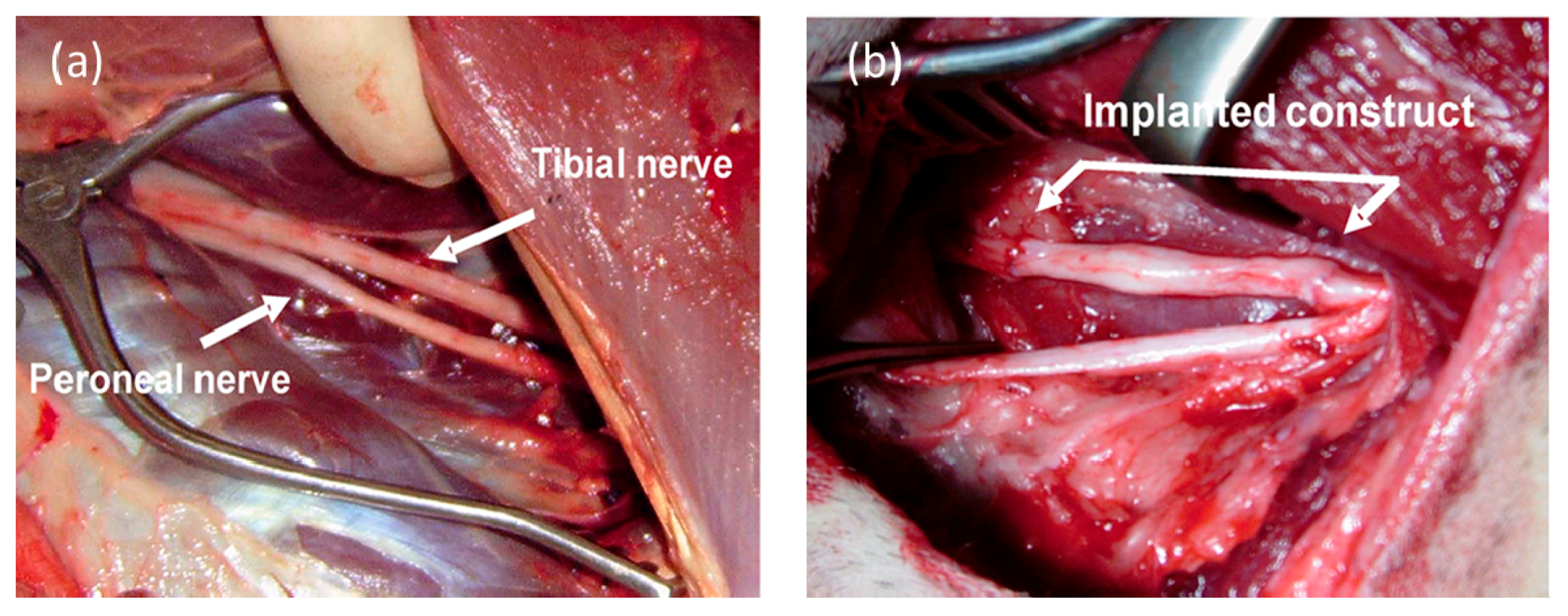{kind=link}
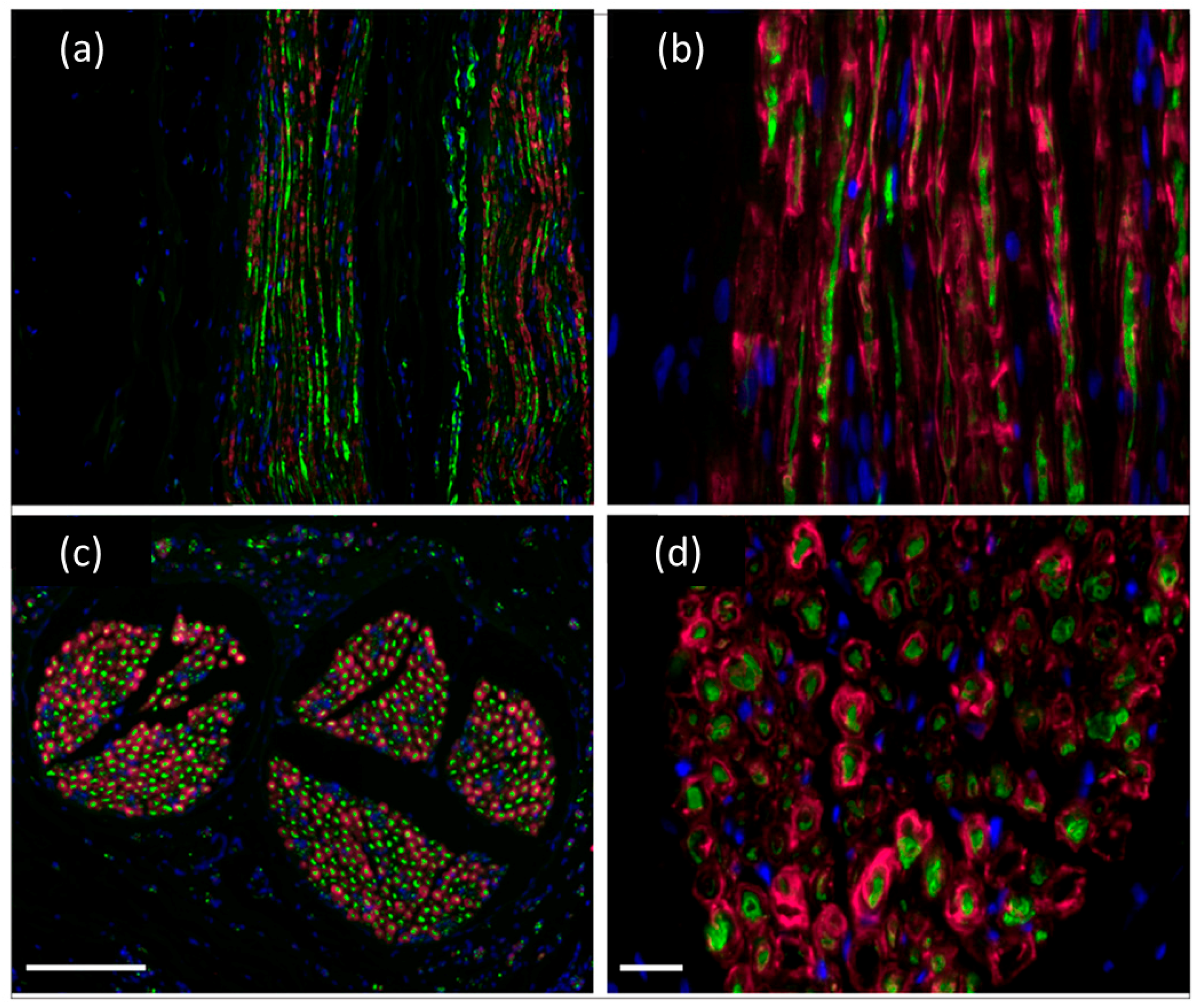{kind=link}
Abstract
:1. Introduction
2. Peripheral Nerve Repair by Nerve Grafting or Conduit Implantation
3. Natural Silk and Silk Analogue-Based Guidance Conduits for Peripheral Nerve Regeneration
3.1. Silk-Neuronal Compatibility
3.2. Biodegradability of Silk
4. The Development of a Spider Silk/Acellularized Vein-Based Nerve Conduit
5. Application of the Spider Silk Conduit for Nerve Repair in Small and Large Animal Models
6. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| DRG | Dorsal root ganglia |
| hMSCs | Human mesenchymal stem cells |
| NGF | nerve growth factor |
| NIH-3T3 | murine fibroblasts |
| Ntera | Human model neurons |
| PGA | polyglycolic acid |
| PHB | poly-3-hydroxybutyrate |
| PLLA | poly l-lactic acid Three letter acronym |
References
- Altman, G.H.; Diaz, F.; Jakuba, C.; Calabro, T.; Horan, R.L.; Chen, J.; Lu, H.; Richmond, J.; Kaplan, D.L. Silk-based biomaterials. Biomaterials 2003, 24, 401–416. [Google Scholar] [CrossRef]
- Yonemura, N.; Mita, K.; Tamura, T.; Sehnal, F. Conservation of silk genes in Trichoptera and Lepidoptera. J. Mol. Evol. 2009, 68, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Craig, C.L. Evolution of arthropod silks. Annu. Rev. Entomol. 1997, 42, 231–267. [Google Scholar] [CrossRef] [PubMed]
- Craig, C.L.; Hsu, M.; Kaplan, D.; Pierce, N.E. A comparison of the composition of silk proteins produced by spiders and insects. Int. J. Biol. Macromol. 1999, 24, 109–118. [Google Scholar] [CrossRef]
- Heim, M.; Keerl, D.; Scheibel, T. Spider silk: From soluble protein to extraordinary fiber. Angew. Chem. Int. Ed. 2009, 48, 3584–3596. [Google Scholar] [CrossRef] [PubMed]
- Lefèvre, T.; Boudreault, S.; Cloutier, C.; Pézolet, M. Diversity of molecular transformations involved in the formation of spider silks. J. Mol. Biol. 2011, 405, 238–253. [Google Scholar]
- Cao, Y.; Wang, B. Biodegradation of silk biomaterials. Int. J. Mol. Sci. 2009, 10, 1514–1524. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Yu, H.S.; Shen, Y.H.; Banno, Y.; Xiang, Z.H.; Zhang, Z. Phylogeny and evolutionary history of the silkworm. Sci. China Life Sci. 2012, 55, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.; Newman, C. Oh what a tangled web: The medicinal uses oe spider silk. Int. J. Dermatol. 1995, 34, 290–292. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rigueiro, J.; Elices, M.; Plaza, G.; Real, J.I.; Guinea, G.V. The effect of spinning forces on spider silk properties. J. Exp. Biol. 2005, 208, 2633–2639. [Google Scholar] [CrossRef] [PubMed]
- Gatesy, J.; Hayashi, C.; Motriuk, D.; Woods, J.; Lewis, R. Extreme diversity, conservation, and convergence of spider silk fibroin sequences. Science 2001, 291, 2603–2605. [Google Scholar] [CrossRef] [PubMed]
- Chinali, A.; Vater, W.; Rudakoff, B.; Sponner, A.; Unger, E.; Grosse, F.; Guehrs, K.H.; Weisshart, K. Containment of extended length polymorphisms in silk proteins. J. Mol. Evol. 2010, 70, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.E.; Nelson, S.R.; Jones, J.A.; Koenig, C.; Hinman, M.; Stricker, S.; Lewis, R.V. Distinct contributions of model MaSp1 and MaSp2 like peptides to the mechanical properties of synthetic major ampullate silk fibers as revealed in silico. Nanotechnol. Sci. Appl. 2008, 1, 9–16. [Google Scholar] [PubMed]
- Guan, J.; Vollrath, F.; Porter, D. Two mechanisms for supercontraction in Nephila spider dragline silk. Biomacromolecules 2011, 12, 4030–4035. [Google Scholar] [CrossRef] [PubMed]
- Birch, R. Surgical Disorders of the Peripheral Nerves, 2nd ed.; Springer Science & Business Media: New York, NY, USA, 2011; pp. 123–140. [Google Scholar]
- Seddon, H.J. Nerve grafting. Ann. R. Coll. Surg. Engl. 1963, 32, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Millesi, H. Significance of nerve transplantation in surgery of the peripheral nerves. Zentralbl. Chir. 1975, 100, 1537–1546. [Google Scholar] [PubMed]
- Daoutis, N.K.; Gerostathopoulos, N.E.; Efstathopoulos, D.G.; Misitizis, D.P.; Bouchlis, G.N.; Anagnostou, S.K. Microsurgical reconstruction of large nerve defects using autologous nerve grafts. Microsurgery 1994, 15, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Terzis, J.K. Clinical microsurgery of the peripheral nerve: The state of the art. Clin. Plast. Surg. 1979, 6, 247–267. [Google Scholar] [PubMed]
- Millesi, H. Nerve transplantation for reconstruction of peripheral nerves injured by the use of the microsurgical technic. Minerva. Chir. 1967, 22, 950–951. [Google Scholar] [PubMed]
- Aszmann, O.C.; Korak, K.J.; Luegmair, M.; Frey, M. Bridging critical nerve defects through an acellular homograft seeded with autologous schwann cells obtained from a regeneration neuroma of the proximal stump. J. Reconstr. Microsurg. 2008, 24, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Nectow, A.R.; Marra, K.G.; Kaplan, D.L. Biomaterials for the development of peripheral nerve guidance conduits. Tissue Eng. Part B: Rev. 2012, 18, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Konofaos, P.; ver Halen, J.P. Nerve repair by means of tubulization: Past, present, future. J. Reconstr. Microsurg. 2013, 29, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Mittnacht, U.; Hartmann, H.; Hein, S.; Oliveira, H.; Dong, M.; Pêgo, A.; Kjems, J.; Howard, K.A.; Schlosshauer, B. Chitosan/siRNA nanoparticles biofunctionalize nerve implants and enable neurite outgrowth. Nano Lett. 2010, 10, 3933–3939. [Google Scholar] [CrossRef] [PubMed]
- Haastert-Talini, K.; Geuna, S.; Dahlin, L.B.; Meyer, C.; Stenberg, L.; Freier, T.; Heimann, C.; Barwig, C.; Pinto, L.F.V.; Raimondo, S.; et al. Chitosan tubes of varying degrees of acetylation for bridging peripheral nerve defects. Biomaterials 2013, 34, 9886–9904. [Google Scholar] [CrossRef] [PubMed]
- Evans, G.R.; Brandt, K.; Katz, S.; Chauvin, P.; Otto, L.; Bogle, M.; Wang, B.; Meszlenyi, R.K.; Lu, L.; Mikos, A.G.; et al. Bioactive poly(l-lactic acid) conduits seeded with Schwann cells for peripheral nerve regeneration. Biomaterials 2002, 23, 841–848. [Google Scholar] [CrossRef]
- Matsumoto, K.; Ohnishi, K.; Kiyotani, T.; Sekine, T.; Ueda, H.; Nakamura, T.; Endo, K.; Shimizu, Y. Peripheral nerve regeneration across an 80-mm gap bridged by a polyglycolic acid (PGA)–collagen tube filled with laminin-coated collagen fibers: A histological and electrophysiological evaluation of regenerated nerves. Brain Res. 2000, 868, 315–328. [Google Scholar] [CrossRef]
- Young, R.C.; Wiberg, M.; Terenghi, G. Poly-3-hydroxybutyrate (PHB): A resorbable conduit for long-gap repair in peripheral nerves. Br. J. Plast. Surg. 2002, 55, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Ichihara, S.; Inada, Y.; Nakamura, T. Artificial nerve tubes and their application for repair of peripheral nerve injury: An update of current concepts. Injury 2008, 39, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Meek, M.F.; Coert, J.H. US Food and Drug Administration /Conformit Europe-approved absorbable nerve conduits for clinical repair of peripheral and cranial nerves. Ann. Plast. Surg. 2008, 60, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Liodaki, E.; Bos, I.; Lohmeyer, J.A.; Senyaman, O.; Mauss, K.L.; Siemers, F.; Mailaender, P.; Stang, F. Removal of collagen nerve conduits (NeuraGen) after unsuccessful implantation: Focus on histological findings. J. Reconstr. Microsurg. 2013, 29, 517–521. [Google Scholar] [PubMed]
- Archibald, S.J.; Shefner, J.; Krarup, C.; Madison, R.D. Monkey median nerve repaired by nerve graft or collagen nerve guide tube. J. Neurosci. 1995, 15, 4109–4123. [Google Scholar] [PubMed]
- Lundborg, G.; Rosen, B.; Dahlin, L.; Danielsen, N.; Holmberg, J. Tubular versus conventional repair of median and ulnar nerves in the human forearm: Early results from a prospective, randomized, clinical study. J. Hand Surg. Am. 1997, 22, 99–106. [Google Scholar] [CrossRef]
- Lundborg, G.; Rosén, B.; Dahlin, L.; Holmberg, J.; Rosén, I. Tubular repair of the median or ulnar nerve in the human forearm: A 5-year follow-up. J. Hand Surg. Am. 2004, 29, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Fields, R.D.; Ellisman, M.H. Axons regenerated through silicone tube splices. II. Functional morphology. Exp. Neurol. 1986, 92, 61–74. [Google Scholar] [CrossRef]
- Berrocal, Y.A.; Almeida, V.W.; Levi, A.D. Limitations of nerve repair of segmental defects using acellular conduits. J. Neurosurg. 2013, 119, 733–738. [Google Scholar] [CrossRef] [PubMed]
- De Ruiter, G.C.W.; Malessy, M.J.A.; Yaszemski, M.J.; Windebank, A.J.; Spinner, R.J. Designing ideal conduits for peripheral nerve repair. Neurosurg. Focus 2009, 26. [Google Scholar] [CrossRef] [PubMed]
- Chiono, V.; Tonda-Turo, C.; Ciardelli, G. Artificial Scaffolds for Peripheral Nerve Reconstruction. Int. Rev. Neurobiol. 2009, 87, 173–198. [Google Scholar] [PubMed]
- Sinis, N.; Schaller, H.E.; Schulte-Eversum, C.; Lanaras, T.; Schlosshauer, B.; Doser, M.; Dietz, K.; Rösner, H.; Müller, H.W.; Haerle, M. Comparative neuro tissue engineering using different nerve guide implants. Acta Neurochir. Suppl. 2007, 100, 61–64. [Google Scholar] [PubMed]
- Meek, M.F.; Dijkstra, J.R.; den Dunnen, W.F.A.; Ijkema-Paassen, J.; Schakenraad, J.M.; Gramsbergen, A.; Robinson, P.H. Functional assessment of sciatic nerve reconstruction: Biodegradable poly (DLLA-ε-CL) nerve guides versus autologous nerve grafts. Microsurgery 1999, 19, 381–388. [Google Scholar] [CrossRef]
- Ribeiro-Resende, V.T.; Koenig, B.; Nichterwitz, S.; Oberhoffner, S.; Schlosshauer, B. Strategies for inducing the formation of bands of Büngner in peripheral nerve regeneration. Biomaterials 2009, 30, 5251–5259. [Google Scholar] [CrossRef] [PubMed]
- Taniuchi, M.; Clark, H.B.; Johnson, E.M., Jr. Induction of nerve growth factor receptor in Schwann cells after axotomy. Proc. Natl. Acad. Sci. USA 1986, 83, 4094–4098. [Google Scholar] [CrossRef] [PubMed]
- Radtke, C.; Vogt, P.M.; Devor, M.; Kocsis, J.D. Keratinocytes acting on injured afferents induce extreme neuronal hyperexcitability and chronic pain. Pain 2010, 148, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Kokai, L.E.; Bourbeau, D.; Weber, D.; McAtee, J.; Marra, K.G. Sustained growth factor delivery promotes axonal regeneration in long gap peripheral nerve repair. Tissue Eng. Part A 2011, 17, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- De Boer, R.; Knight, A.M.; Borntraeger, A.; Hebert-Blouin, M.N.; Spinner, R.J.; Malessy, M.J.; Yaszemski, M.J.; Windebank, A.J. Rat sciatic nerve repair with a poly-lactic-co-glycolic acid scaffold and nerve growth factor releasing microspheres. Microsurgery 2011, 31, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Steuer, H.; Fadale, R.; Müller, E.; Müller, H.W.; Planck, H.; Schlosshauer, B. Biohybride nerve guide for regeneration: Degradable polylactide fibers coated with rat Schwann cells. Neurosci. Lett. 1999, 277, 165–168. [Google Scholar] [CrossRef]
- Terenghi, G.; Wiberg, M.; Kingham, P.J. Use of stem cells for improving nerve regeneration. Int. Rev. Neurobiol. 2009, 87, 393–403. [Google Scholar] [PubMed]
- Radtke, C.; Allmeling, C.; Waldmann, K.H.; Reimers, K.; Thies, K.; Schenk, H.C.; Hillmer, A.; Guggenheim, M.; Brandes, G.; Vogt, P.M. Spider silk constructs enhance axonal regeneration and remyelination in long nerve defects in sheep. PLoS ONE 2011, 6, e16990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Chen, X.; Ding, F.; Zhang, P.; Liu, J.; Gu, X. Biocompatibility evaluation of silk fibroin with peripheral nerve tissues and cells in vitro. Biomaterials 2007, 28, 1643–1652. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Ding, F.; Yang, Y.; Hu, N.; Wu, H.; Gu, X. Evaluation on in vitro biocompatibility of silk fibroin-based biomaterials with primarily cultured hippocampal neurons. J. Biomed. Mater. Res. Part A 2009, 91, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Kocsis, J.D.; Honmou, O. Bone marrow stem cells in experimental stroke. Prog. Brain Res. 2012, 201, 79–98. [Google Scholar] [PubMed]
- Garcia-Fuentes, M.; Meinel, A.J.; Hilbe, M.; Meinel, L.; Merkle, H.P. Silk fibroin/hyaluronan scaffolds for human mesenchymal stem cell culture in tissue engineering. Biomaterials 2009, 30, 5068–5076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allmeling, C.; Jokuszies, A.; Reimers, K.; Kall, S.; Vogt, P.M. Use of spider silk fibres as an innovative material in a biocompatible artificial nerve conduit. J. Cell. Mol. Med. 2006, 10, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Allmeling, C.; Jokuszies, A.; Reimers, K.; Kall, S.; Choi, C.Y.; Brandes, G.; Kasper, C.; Scheper, T.; Guggenheim, M.; Vogt, P.M. Spider silk fibres in artificial nerve constructs promote peripheral nerve regeneration. Cell Prolif. 2008, 41, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cai, L. Polymers for fabricating nerve conduits. Int. J. Polym. Sci. 2010, 2010, 1–20. [Google Scholar] [CrossRef]
- Siemionow, M.; Brzezicki, G. Current techniques and concepts in peripheral nerve repair. Int. Rev. Neurobiol. 2009, 87, 141–172. [Google Scholar] [PubMed]
- Hu, X.; Vasanthavada, K.; Kohler, K.; McNary, S.; Moore, A.M.F.; Vierra, C.A. Molecular mechanisms of spider silk. Cell. Mol. Life Sci. 2006, 63, 1986–1999. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.S.; Soni, S.; Le, C.; Biasca, M.; Farr, E.; Chen, E.Y.T.; Chin, W.C. Human stem cell neuronal differentiation on silk-carbon nanotube composite. Nanoscale Res. Lett. 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ding, F.; Wu, J.; Hu, W.; Liu, W.; Liu, J.; Gu, X. Development and evaluation of silk fibroin-based nerve grafts used for peripheral nerve regeneration. Biomaterials 2007, 28, 5526–5535. [Google Scholar] [CrossRef] [PubMed]
- Hu, A.; Zuo, B.; Zhang, F.; Zhang, H.; Lan, Q. Evaluation of electronspun silk fibroin-based transplants used for facial nerve repair. Otol. Neurotol. 2013, 23, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Kuhbier, J.W.; Reimers, K.; Kasper, C.; Allmeling, C.; Hillmer, A.; Menger, B.; Vogt, P.M.; Radtke, C. First investigation of spider silk as a braided microsurgical suture. J. Biomed. Mater. Res. Part B Appl. Biomater. 2011, 97, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Strauß, S.; Neumeister, A.; Barcikowski, S.; Kracht, D.; Kuhbier, J.W.; Radtke, C.; Reimers, K.; Vogt, P.M. Adhesion, vitality and osteogenic differentiation capacity of adipose derived stem cells seeded on nitinol nanoparticle coatings. PLoS ONE 2013, 8, e53309. [Google Scholar] [CrossRef] [PubMed]
- Wendt, H.; Hillmer, A.; Reimers, K.; Kuhbier, J.W.; Schäfer-Nolte, F.; Allmeling, C.; Kasper, C.; Vogt, P.M. Artificial skin-culturing of different skin cell lines for generating an artificial skin substitute on cross-weaved spider silk fibres. PLoS ONE 2011, 6, e21833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfer-Nolte, F.; Hennecke, K.; Reimers, K.; Schnabel, R.; Allmeling, C.; Vogt, P.M.; Kuhbier, J.W.; Mirastschijski, U. Biomechanics and biocompatibility of woven spider silk meshes during remodeling in a rodent fascia replacement model. Ann. Surg. 2014, 259, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Hennecke, K.; Redeker, J.; Kuhbier, J.W.; Strauss, S.; Allmeling, C.; Kasper, C.; Reimers, K.; Vogt, P.M. Bundles of spider silk, braided into sutures, resist basic cyclic tests: potential use for flexor tendon repair. PLoS ONE 2013, 8, e61100. [Google Scholar] [CrossRef] [PubMed]
- Kuhbier, J.W.; Allmeling, C.; Reimers, K.; Hillmer, A.; Kasper, C.; Menger, B.; Brandes, G.; Guggenheim, M.; Vogt, P.M. Interactions between spider silk and cells–NIH/3T3 fibroblasts seeded on miniature weaving frames. PLoS ONE 2010, 5, e12032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roloff, F.; Strauß, S.; Vogt, P.M.; Bicker, G.; Radtke, C. Spider silk as guiding biomaterial for human model neurons. BioMed Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Vollrath, F. Surprising strength of silkworm silk. J. Appl. Polym. Sci. 2001, 82, 1928–1935. [Google Scholar]
- Lazaris, A.; Arcidiacono, S.; Huang, Y.; Zhou, J.F.; Duguay, F.; Chretien, N.; Welsh, E.A.; Soares, J.W.; Karatzas, C.N. Spider silk fibers spun from soluble recombinant silk produced in mammalian cells. Science 2002, 295, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Cunniff, P.M.; Fossey, S.A.; Auerbach, M.A.; Song, J.W. Mechanical properties of mahor ampulate gland silk fibers extracted from Nephila clavipes spiders. Silk Polym. 1993, 544, 234–251. [Google Scholar]
- Radtke, C.; Kocsis, J.D.; Reimers, K.; Allmeling, C.; Vogt, P.M. Sural nerve defects after nerve biopsy or nerve transfer as a sensory regeneration model for peripheral nerve conduit implantation. Med. Hypotheses 2013, 81, 500–502. [Google Scholar] [CrossRef] [PubMed]


© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radtke, C. Natural Occurring Silks and Their Analogues as Materials for Nerve Conduits. Int. J. Mol. Sci. 2016, 17, 1754. https://doi.org/10.3390/ijms17101754
Radtke C. Natural Occurring Silks and Their Analogues as Materials for Nerve Conduits. International Journal of Molecular Sciences. 2016; 17(10):1754. https://doi.org/10.3390/ijms17101754
Chicago/Turabian StyleRadtke, Christine. 2016. "Natural Occurring Silks and Their Analogues as Materials for Nerve Conduits" International Journal of Molecular Sciences 17, no. 10: 1754. https://doi.org/10.3390/ijms17101754