Aberrant Expression of Posterior HOX Genes in Well Differentiated Histotypes of Thyroid Cancers


Abstract
:1. Introduction
2. Results
2.1. Clinic Pathological Characteristics of Thyroid Cancers and Patients
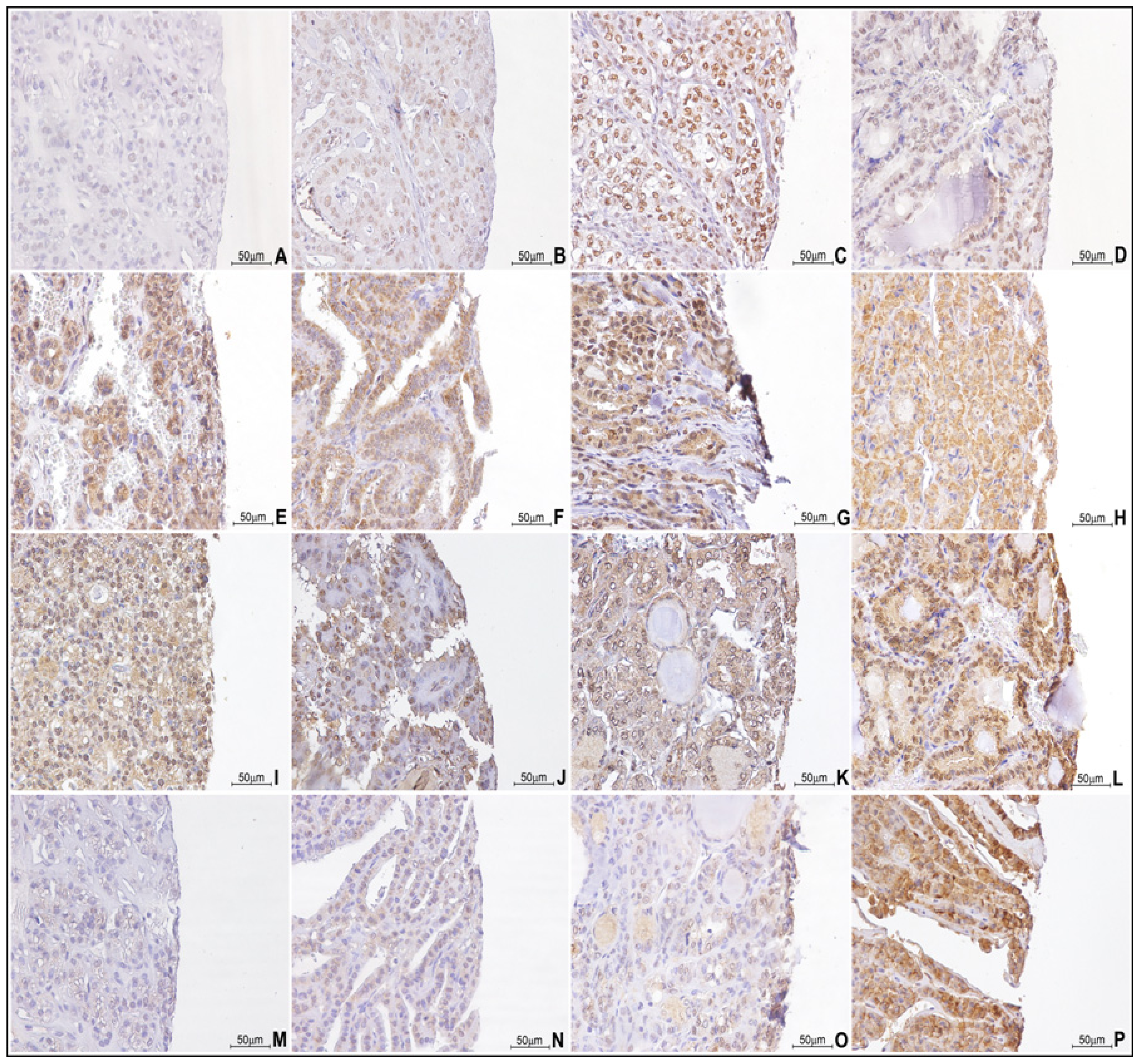
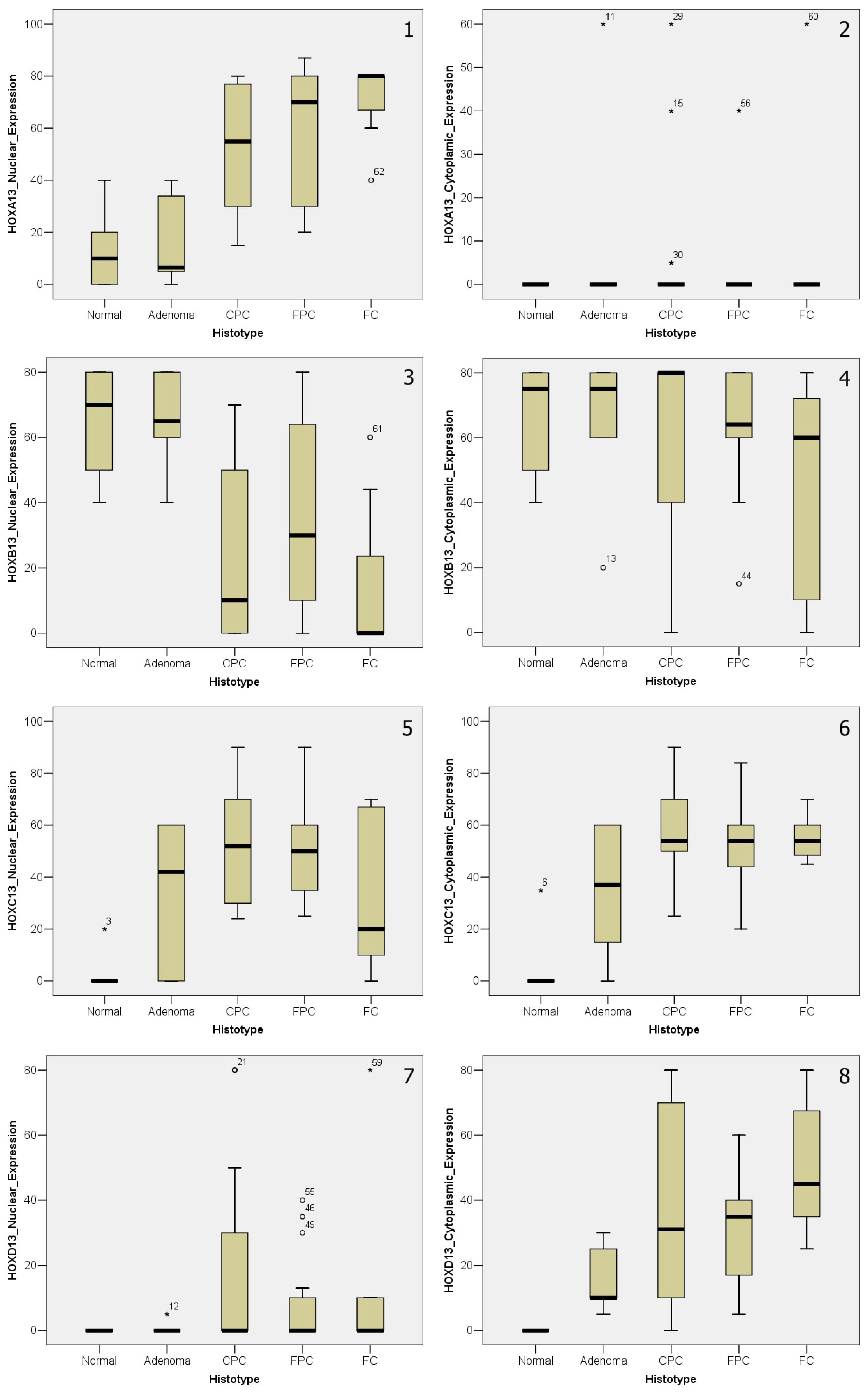
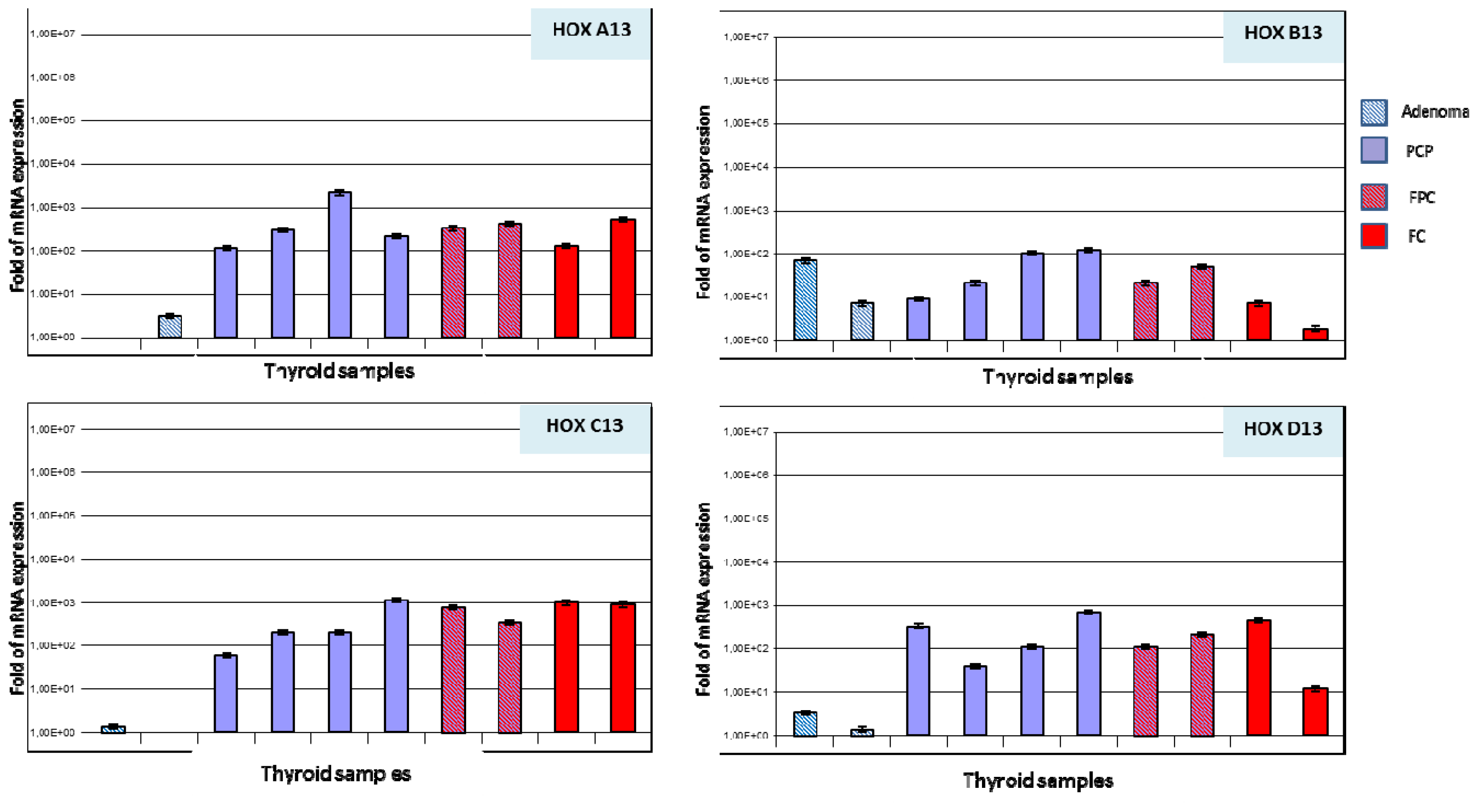
2.2. Paralogous 13 HOX Genes Expression in Thyroid Cancer Histotypes
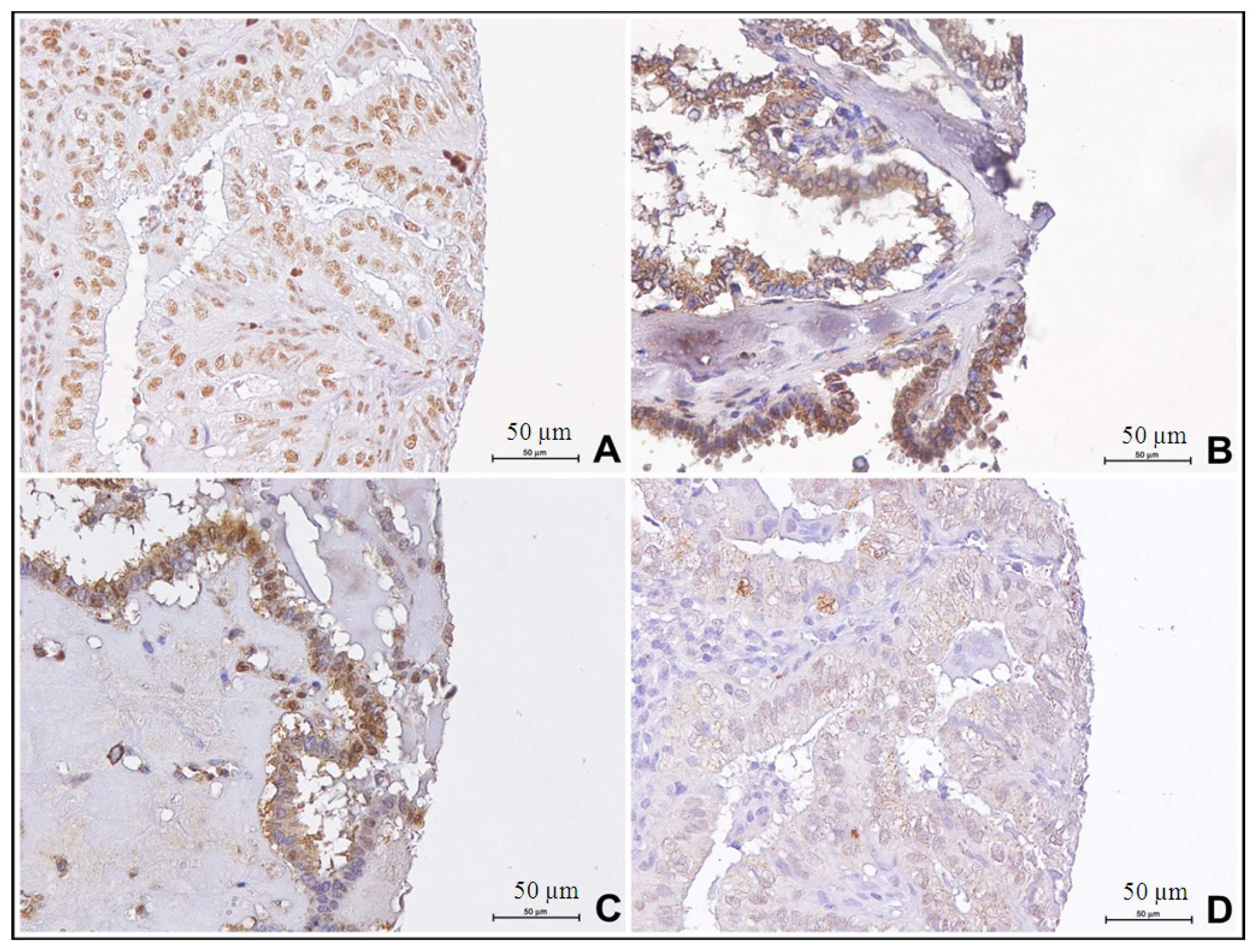
2.3. Paralogous 13 HOX Genes Expression in Papillary and Follicular Thyroid Cancer Progression
3. Discussion
4. Methods
4.1. Thyroid Cancer Patients
4.2. TMAs Building
4.3. Immunohistochemistry Analysis
4.4. Evaluation of Immunostaining
4.5. RNA Extraction and Analysis
4.6. Quantitative Real-Time PCR
4.7. Statistical Analysis
5. Conclusions
Supplementary Information
ijms-14-21727-s001.pdf




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gender | M | 12 (24) |
| F | 38 (76) | |
| Age | <=45 | 29 (58) |
| 46+ | 21 (42) | |
| HISTOTYPE | CPC | 26 (52) |
| FPC | 17 (34) | |
| FC | 7 (14) | |
| STAGE | I | 29 (58) |
| II | 2 (4) | |
| III | 13 (26) | |
| IV | 6 (12) | |
| Wilcoxon Signed Ranks Test | ||
|---|---|---|
| Ranks | N | p-Value |
| HOX A13 | ||
| Cytoplamic_Expression < Nuclear_Expression | 57 | <0.001 |
| Cytoplamic_Expression > Nuclear_Expression | 1 | |
| Cytoplamic_Expression = Nuclear_Expression | 4 | |
| HOX B13 | ||
| Cytoplasmic_Expression < Nuclear_Expression | 3 | <0.001 |
| Cytoplasmic_Expression > Nuclear_Expression | 47 | |
| Cytoplasmic_Expression = Nuclear_Expression | 12 | |
| HOX C13 | ||
| Cytoplasmic_Expression < Nuclear_Expression | 21 | 0.19 |
| Cytoplasmic_Expression > Nuclear_Expression | 29 | |
| Cytoplasmic_Expression = Nuclear_Expression | 13 | |
| HOX D13 | ||
| Cytoplasmic_Expression < Nuclear_Expression | 4 | <0.001 |
| Cytoplasmic_Expression > Nuclear_Expression | 50 | |
| Cytoplasmic_Expression = Nuclear_Expression | 9 | |
Conflicts of Interest
References
- Sherman, S.I. Thyroid carcinoma. Lancet 2003, 361, 501–511. [Google Scholar]
- Aldred, M.A.; Huang, Y.; Liyanarachchi, S.; Pellegata, N.S.; Gimm, O.; Jhiang, S.; Davuluri, R.V.; de la Chapelle, A.; Eng, C. Papillary and follicular thyroid carcinomas show distinctly different microarray expression profiles and can be distinguished by a minimum of five genes. J. Clin. Oncol 2004, 22, 3531–3539. [Google Scholar]
- Kouniavsky, G.; Zeiger, M.A. Thyroid tumorigenesis and molecular markers in thyroid cancer. Curr. Opin. Oncol 2010, 22, 23–29. [Google Scholar]
- Santoro, M.; Melillo, R.M.; Fusco, A. RET/PTC activation in papillary thyroid carcinoma: European Journal of Endocrinology Prize Lecture. Eur. J. Endocrinol 2006, 155, 645–653. [Google Scholar]
- Leonardi, G.C.; Candido, S.; Carbone, M.; Raiti, F.; Colaianni, V.; Garozzo, S.; Cinà, D.; McCubrey, J.A.; Libra, M. BRAF mutations in papillary thyroid carcinoma and emerging targeted therapies. Mol. Med. Rep 2012, 6, 687–694. [Google Scholar]
- Cillo, C. HOX genes in human cancers. Invas. Metastasis 1994–1995, 14, 38–49. [Google Scholar]
- Cillo, C.; Faiella, A.; Cantile, M.; Boncinelli, E. Homeobox genes and cancer. Exp. Cell Res 1999, 248, 1–9. [Google Scholar]
- Nunes, F.D.; de Almeida, F.C.; Tucci, R.; de Sousa, S.C. Homeobox genes: A molecular link between development and cancer. Pesqui. Odontol. Bras 2003, 17, 94–98. [Google Scholar]
- Grier, D.G.; Thompson, A.; Kwasniewska, A.; McGonigle, G.J.; Halliday, H.L.; Lappin, T.R. The pathophysiology of HOX genes and their role in cancer. J. Pathol 2005, 205, 154–171. [Google Scholar]
- Argiropoulos, B.; Humphries, R.K. Hox genes in hematopoiesis and leukemogenesis. Oncogene 2007, 26, 6766–6776. [Google Scholar]
- Shah, N.; Sukumar, S. The Hox genes and their roles in oncogenesis. Nat. Rev. Cancer 2010, 10, 361–371. [Google Scholar]
- Cantile, M.; Franco, R.; Tschan, A.; Baumhoer, D.; Zlobec, I.; Schiavo, G.; Forte, I.; Bihl, M.; Liguori, G.; Botti, G.; et al. HOX D13 expression across 79 tumor tissue types. Int. J. Cancer 2009, 125, 1532–1541. [Google Scholar] [Green Version]
- Cillo, C.; Schiavo, G.; Cantile, M.; Bihl, M.P.; Sorrentino, P.; Carafa, V.; D’Armiento, M.; Roncalli, M.; Sansano, S.; Vecchione, R.; et al. The HOX gene network in hepatocellular carcinoma. Int. J. Cancer 2011, 129, 2577–2587. [Google Scholar]
- Simon, R.; Struckmann, K.; Schraml, P.; Wagner, U.; Forster, T.; Moch, H.; Fijan, A.; Bruderer, J.; Wilber, K.; Mihatsch, M.J.; et al. Amplification pattern of 12q13–q15 genes (MDM2, CDK4, GLI) in urinary bladder cancer. Oncogene 2002, 21, 2476–2483. [Google Scholar]
- Wikman, H.; Nymark, P.; Väyrynen, A.; Jarmalaite, S.; Kallioniemi, A.; Salmenkivi, K.; Vainio-Siukola, K.; Husgafvel-Pursiainen, K.; Knuutila, S.; Wolf, M.; et al. CDK4 is a probable target gene in a novel amplicon at 12q13.3–q14.1 in lung cancer. Genes Chrom. Cancer 2005, 42, 193–199. [Google Scholar]
- Muthusamy, V.; Hobbs, C.; Nogueira, C.; Cordon-Cardo, C.; McKee, P.H.; Chin, L.; Bosenberg, M.W. Amplification of CDK4 and MDM2 in malignant melanoma. Genes Chrom. Cancer 2006, 45, 447–454. [Google Scholar]
- Willmore-Payne, C.; Holden, J.; Turner, K.C.; Proia, A.; Layfield, L.J. Translocations and amplifications of chromosome 12 in liposarcoma demonstrated by the LSI CHOP breakapart rearrangement probe. Arch. Pathol. Lab. Med 2008, 132, 952–957. [Google Scholar]
- Barr, F.G.; Duan, F.; Smith, L.M.; Gustafson, D.; Pitts, M.; Hammond, S.; Gastier-Foster, J.M. Genomic and clinical analyses of 2p24 and 12q13–q14 amplification in alveolar rhabdomyosarcoma: A report from the children’s oncology group. Genes Chrom. Cancer 2009, 48, 661–672. [Google Scholar]
- Or, Y.Y.; Chung, G.T.; To, K.F.; Chow, C.; Choy, K.W.; Tong, C.Y.; Leung, A.W.; Hui, A.B.; Tsao, S.W.; Ng, H.K.; et al. Identification of a novel 12p13.3 amplicon in nasopharyngeal carcinoma. J. Pathol 2010, 220, 97–107. [Google Scholar]
- Fischer, U.; Leidinger, P.; Keller, A.; Folarin, A.; Ketter, R.; Graf, N.; Lenhof, H.P.; Meese, E. Amplicons on chromosome 12q13–21 in glioblastoma recurrences. Int. J. Cancer 2010, 126, 2594–2602. [Google Scholar]
- Mejia-Guerrero, S.; Quejada, M.; Gokgoz, N.; Gill, M.; Parkes, R.K.; Wunder, J.S.; Andrulis, I.L. Characterization of the 12q15 MDM2 and 12q13–14 CDK4 amplicons and clinical correlations in osteosarcoma. Genes Chrom. Cancer 2010, 49, 518–525. [Google Scholar]
- Cantile, M.; Scognamiglio, G.; Anniciello, A.; Farina, M.; Gentilcore, G.; Santonastaso, C.; Fulciniti, F.; Cillo, C.; Franco, R.; Ascierto, P.A.; et al. Increased HOX C13 expression in metastatic melanoma progression. J. Transl. Med 2012, 10, 91. [Google Scholar]
- Marra, L.; Cantile, M.; Scognamiglio, G.; Perdonà, S.; la Mantia, E.; Cerrone, M.; Gigantino, V.; Cillo, C.; Caraglia, M.; Pignata, S.; et al. Deregulation of HOX B13 expression in urinary bladder cancer progression. Curr. Med. Chem 2013, 20, 833–839. [Google Scholar]
- Cantile, M.; Pettinato, G.; Procino, A.; Feliciello, I.; Cindolo, L.; Cillo, C. In vivo expression of the whole HOX gene network in huma breast cancer. Eur. J. Cancer 2003, 39, 257–264. [Google Scholar]
- Miao, J.; Wang, Z.; Provencher, H.; Muir, B.; Dahiya, S.; Carney, E.; Leong, C.O.; Sgroi, D.C.; Orsulic, S. HOX B13 promotes ovarian cancer progression. Proc. Natl. Acad. Sci. USA 2007, 104, 17093–17098. [Google Scholar]
- Muragaki, Y.; Mundlos, S.; Upton, J.; Olsen, B.R. Altered growth and branching patterns in synpolydactyly caused by mutations in HOX D13. Science 1996, 272, 548–551. [Google Scholar]
- Mortlock, D.P.; Innis, J. Mutation of HOX A13 in hand-foot-genital syndrome. Nat. Genet 1997, 15, 179–180. [Google Scholar]
- Gu, Z.D.; Shen, L.Y.; Wang, H.; Chen, X.M.; Li, Y.; Ning, T.; Chen, K.N. HOX A13 promotes cancer cell growth and predicts poor survival of patients with esophageal squamous cell carcinoma. Cancer Res 2009, 69, 4969–4973. [Google Scholar]
- Han, Y.; Tu, W.W.; Wen, Y.G.; Li, D.P.; Qiu, G.Q.; Tang, H.M.; Peng, Z.H.; Zhou, C.Z. Identification and validation that up-expression of HOX A13 is a novel independent prognostic marker of a worse outcome in gastric cancer based on immunohistochemistry. Med. Oncol 2013, 30, 564. [Google Scholar]
- Ma, X.J.; Wang, Z.; Ryan, P.D.; Isakoff, S.J.; Barmettler, A.; Fuller, A.; Muir, B.; Mohapatra, G.; Salunga, R.; Tuggle, J.T.; et al. A two-gene expression ratio predicts clinical outcome in breast cancer patients treated with tamoxifen. Cancer Cell 2004, 5, 607–616. [Google Scholar]
- McMullin, R.P.; Mutton, L.N.; Bieberich, C.J. Hoxb13 regulatory elements mediate transgene expression during prostate organogenesis and carcinogenesis. Dev. Dyn 2009, 238, 664–672. [Google Scholar]
- Akbari, M.R.; Trachtenberg, J.; Lee, J.; Tam, S.; Bristow, R.; Loblaw, A.; Narod, S.A.; Nam, R.K. Association between germline HOXB13 G84E mutation and risk of prostate cancer. J. Natl. Cancer Inst 2012, 104, 1260–1262. [Google Scholar]
- Maeda, K.; Hamada, J.; Takahashi, Y.; Tada, M.; Yamamoto, Y.; Sugihara, T.; Moriuchi, T. Altered expressions of HOX genes in human cutaneous malignant melanoma. Int. J. Cancer 2005, 114, 436–441. [Google Scholar]
- López, R.; Garrido, E.; Piña, P.; Hidalgo, A.; Lazos, M.; Ochoa, R.; Salcedo, M. HOXB homeobox gene expression in cervical carcinoma. Int. J. Gynecol. Cancer 2006, 16, 329–335. [Google Scholar]
- Jung, C.; Kim, R.S.; Zhang, H.; Lee, S.J.; Sheng, H.; Loehrer, P.J.; Gardner, T.A.; Jeng, M.H.; Kao, C. HOXB13 is downregulated in colorectal cancer to confer TCF4-mediated transactivation. Br. J. Cancer 2005, 92, 2233–2239. [Google Scholar]
- Cantile, M.; Cindolo, L.; Napodano, G.; Altieri, V.; Cillo, C. Hyperexpression of locus C genes in the HOX network is strongly associated in vivo with human bladder transitional cell carcinomas. Oncogene 2003, 22, 6462–6468. [Google Scholar]
- Cantile, M.; Schiavo, G.; Franco, R.; Cindolo, L.; Procino, A.; D’Armiento, M.; Facchini, G.; Terracciano, L.; Botti, G.; Cillo, C. Expression of lumbosacral HOX genes, crucial in kidney organogenesis, is systematically deregulated in clear cell kidney cancers. Anticancer Drugs 2011, 22, 392–401. [Google Scholar]
- Takahashi, Y.; Hamada, J.; Murakawa, K.; Takada, M.; Tada, M.; Nogami, I.; Hayashi, N.; Nakamori, S.; Monden, M.; Miyamoto, M.; et al. Expression profiles of 39 HOX genes in normal human adult organs and anaplastic thyroid cancer cell lines by quantitative real-time RT-PCR system. Exp. Cell Res 2004, 293, 144–153. [Google Scholar]
- Wiseman, S.M.; Loree, T.R.; Rigual, N.R.; Hicks, W.L., Jr.; Winston, J.S.; Swede, H.; Bartos, J.D.; Anderson, G.R.; Stoler, D.L. Papillary thyroid cancer: High inter-(simple sequence repeat) genomic instability in a typically indolent cancer. Head Neck 2003, 25, 825–832. [Google Scholar]
- Gandhi, M.; Evdokimova, V.; Nikiforov, Y.E. Mechanisms of chromosomal rearrangements in solid tumors: The model of papillary thyroid carcinoma. Mol. Cell Endocrinol 2010, 321, 36–43. [Google Scholar]
- Giordano, T.J.; Kuick, R.; Thomas, D.G.; Misek, D.E.; Vinco, M.; Sanders, D.; Zhu, Z.; Ciampi, R.; Roh, M.; Shedden, K.; et al. Molecular classification of papillary thyroid carcinoma: Distinct BRAF, RAS, and RET/PTC mutation-specific gene expression profiles discovered by DNA microarray analysis. Oncogene 2005, 24, 6646–6656. [Google Scholar]
- Bychkov, A.; Saenko, V.; Nakashima, M.; Mitsutake, N.; Rogounovitch, T.; Nikitski, A.; Orim, F.; Yamashita, S. Patterns of FOXE1 expression in papillary thyroid carcinoma by immunohistochemistry. Thyroid 2013, 23, 817–828. [Google Scholar]
- Sancisi, V.; Borettini, G.; Maramotti, S.; Ragazzi, M.; Tamagnini, I.; Nicoli, D.; Piana, S.; Ciarrocchi, A. Runx2 isoform I controls a panel of proinvasive genes driving aggressiveness of papillary thyroid carcinomas. J. Clin. Endocrinol. Metable 2012, 97, E2006–E2015. [Google Scholar]
- Gilbert-Sirieix, M.; Makoukji, J.; Kimura, S.; Talbot, M.; Caillou, B.; Massaad, C.; Massaad-Massade, L. Wnt/β-catenin signaling pathway is a direct enhancer of thyroid transcription factor-1 in human papillary thyroid carcinoma cells. PLoS One 2011, 6, e22280. [Google Scholar]
- Kömüves, L.G.; Ma, X.K.; Stelnicki, E.; Rozenfeld, S.; Oda, Y.; Largman, C. HOXB13 homeodomain protein is cytoplasmic throughout fetal skin development. Dev. Dyn 2003, 227, 192–202. [Google Scholar]
- Chile, T.; Fortes, M.A.; Corrêa-Giannella, M.L.; Brentani, H.P.; Augusto, D.M.; Puga, R.D.; de Paula, V.D.; Kubrusly, M.S.; Novak, E.M.; Bacchella, T.; et al. HOXB7 mRNA is overexpressed in pancreatic ductal adenocarcinomas and its knockdown induces cell cycle arrest and apoptosis. BMC Cancer 2013, 3, 451. [Google Scholar]
- Morgan, R.; Pirard, P.M.; Shears, L.; Sohal, J.; Pettengell, R.; Pandha, H.S. Antagonism of HOX/PBX dimer formation blocks the in vivo proliferation of melanoma. Cancer Res 2007, 67, 5806–5813. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cantile, M.; Scognamiglio, G.; La Sala, L.; La Mantia, E.; Scaramuzza, V.; Valentino, E.; Tatangelo, F.; Losito, S.; Pezzullo, L.; Chiofalo, M.G.; et al. Aberrant Expression of Posterior HOX Genes in Well Differentiated Histotypes of Thyroid Cancers. Int. J. Mol. Sci. 2013, 14, 21727-21740. https://doi.org/10.3390/ijms141121727
Cantile M, Scognamiglio G, La Sala L, La Mantia E, Scaramuzza V, Valentino E, Tatangelo F, Losito S, Pezzullo L, Chiofalo MG, et al. Aberrant Expression of Posterior HOX Genes in Well Differentiated Histotypes of Thyroid Cancers. International Journal of Molecular Sciences. 2013; 14(11):21727-21740. https://doi.org/10.3390/ijms141121727
Chicago/Turabian StyleCantile, Monica, Giosuè Scognamiglio, Lucia La Sala, Elvira La Mantia, Veronica Scaramuzza, Elena Valentino, Fabiana Tatangelo, Simona Losito, Luciano Pezzullo, Maria Grazia Chiofalo, and et al. 2013. "Aberrant Expression of Posterior HOX Genes in Well Differentiated Histotypes of Thyroid Cancers" International Journal of Molecular Sciences 14, no. 11: 21727-21740. https://doi.org/10.3390/ijms141121727