Dual PI3K/mTOR Inhibitor NVP-BEZ235 Enhances Radiosensitivity of Head and Neck Squamous Cell Carcinoma (HNSCC) Cell Lines Due to Suppressed Double-Strand Break (DSB) Repair by Non-Homologous End Joining

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
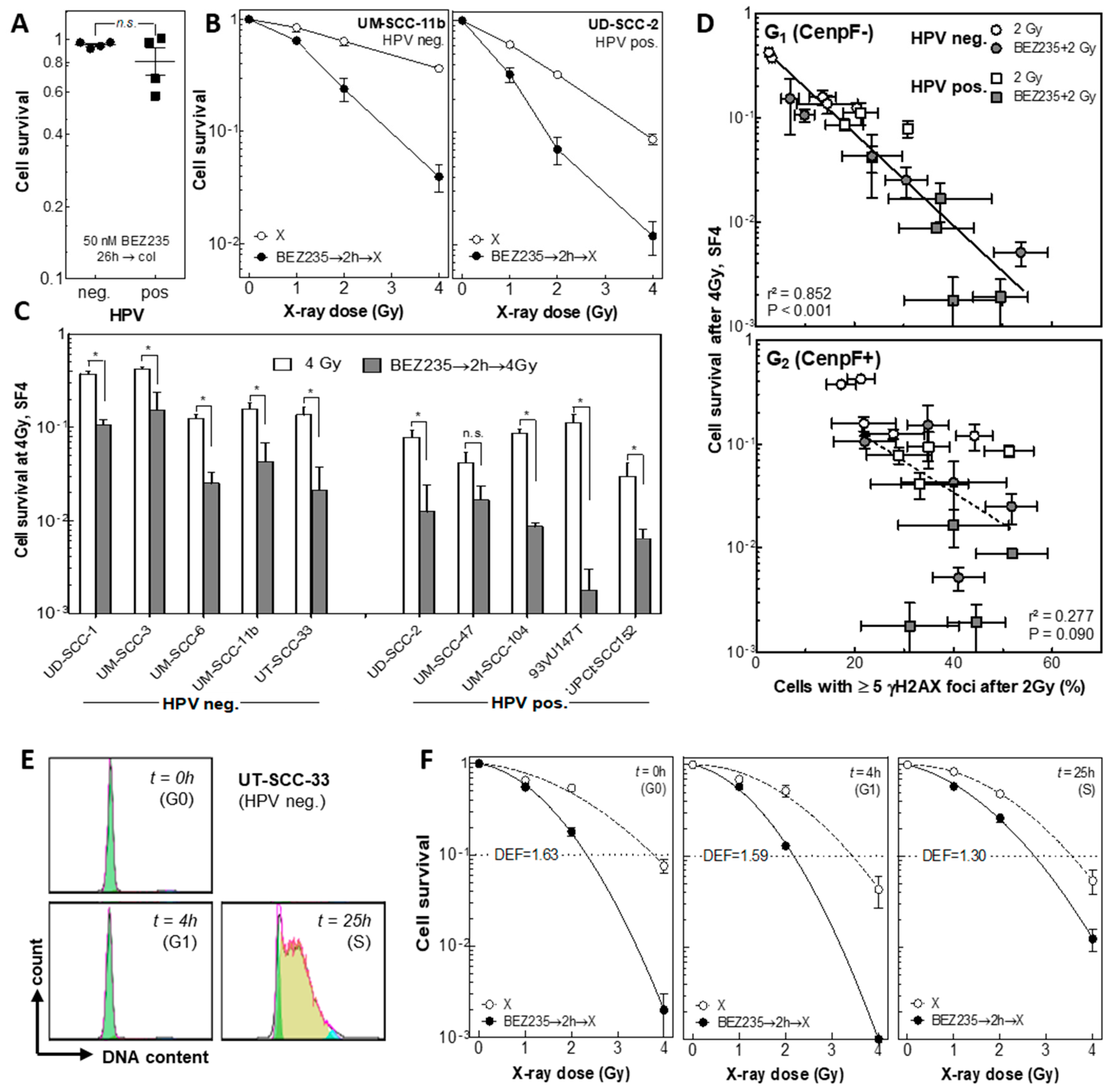
2. Results
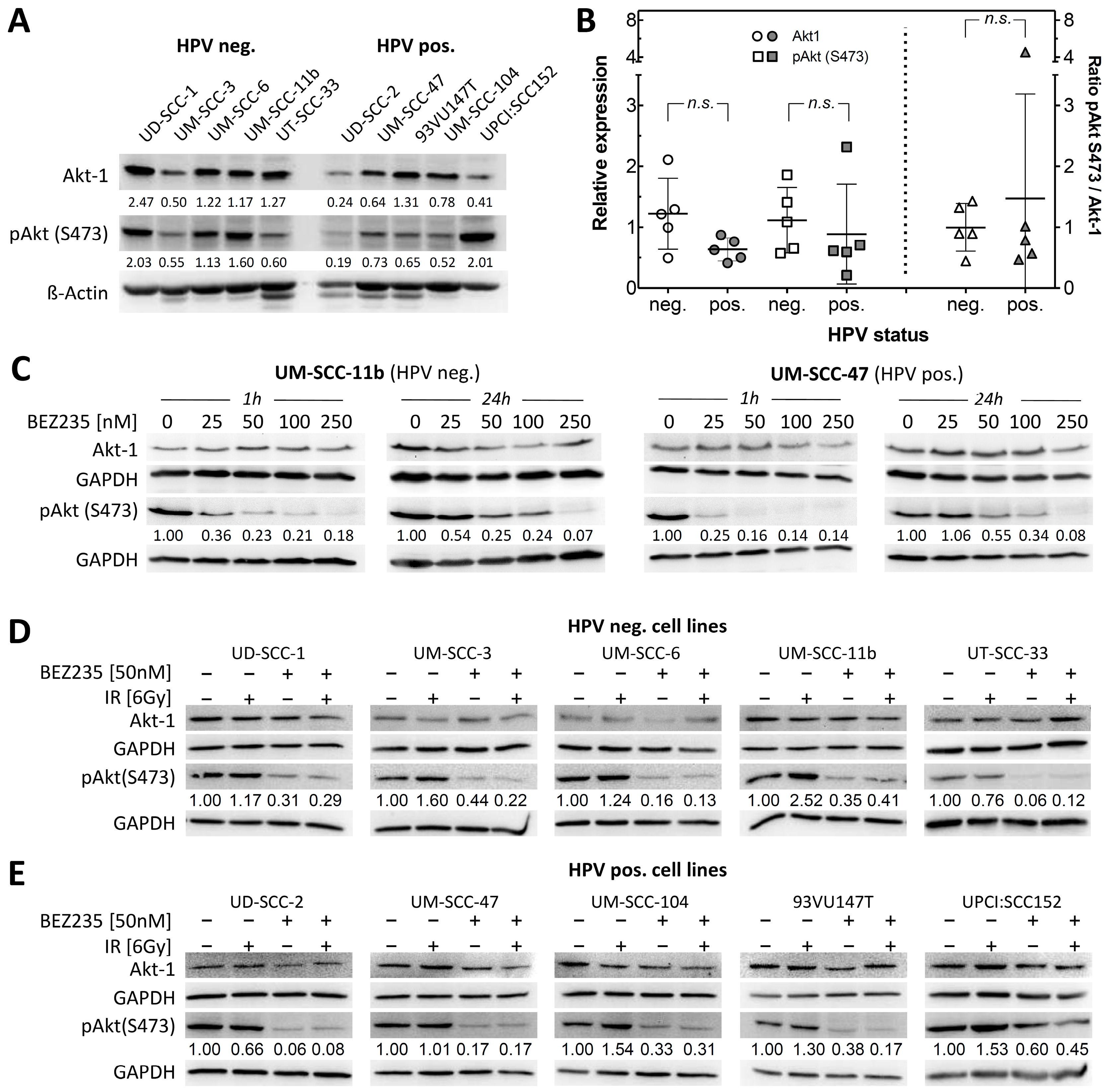
2.1. Akt Status in HPV Negative and Positive HNSCC Cells Shows Similar Variation
2.2. BEZ235 Suppresses Akt Phosphorylation on Ser473 Independent of HPV Status
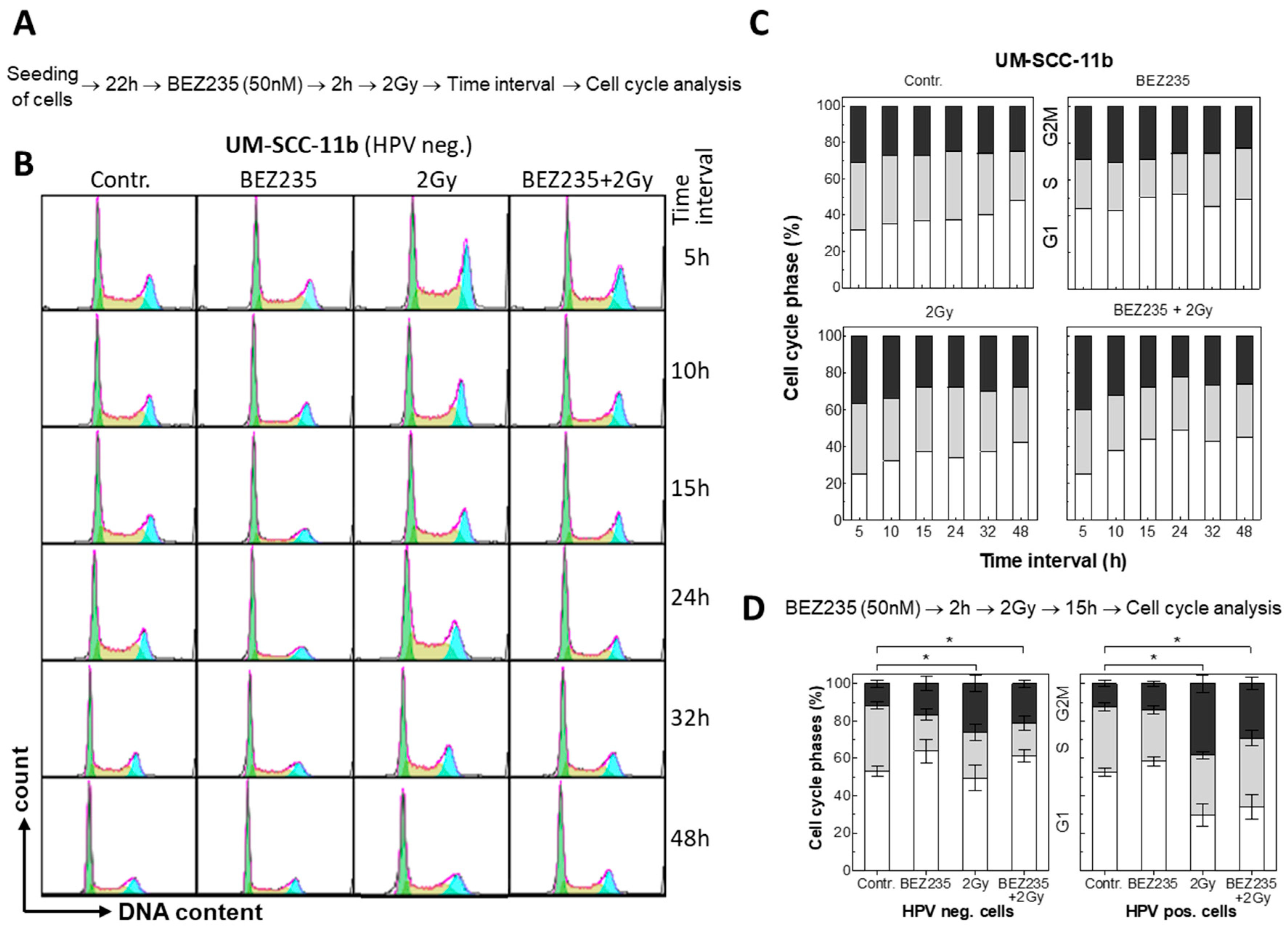
2.3. BEZ235 Leads to a Moderate G1 Arrest without Affecting Radiation-Induced Apoptosis
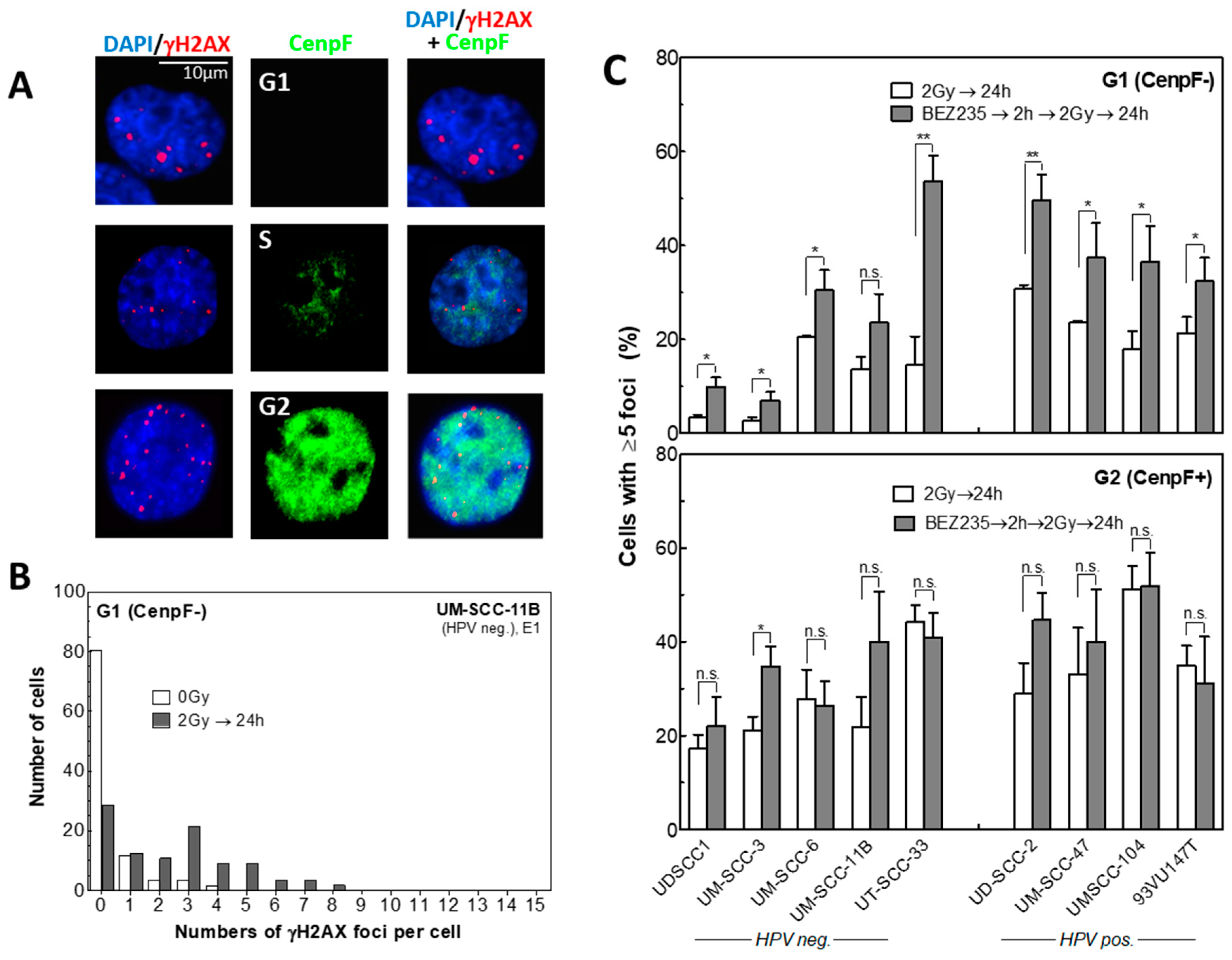
2.4. BEZ235 Strongly Inhibits the Repair of Radiation-Induced DSBs Only in G1
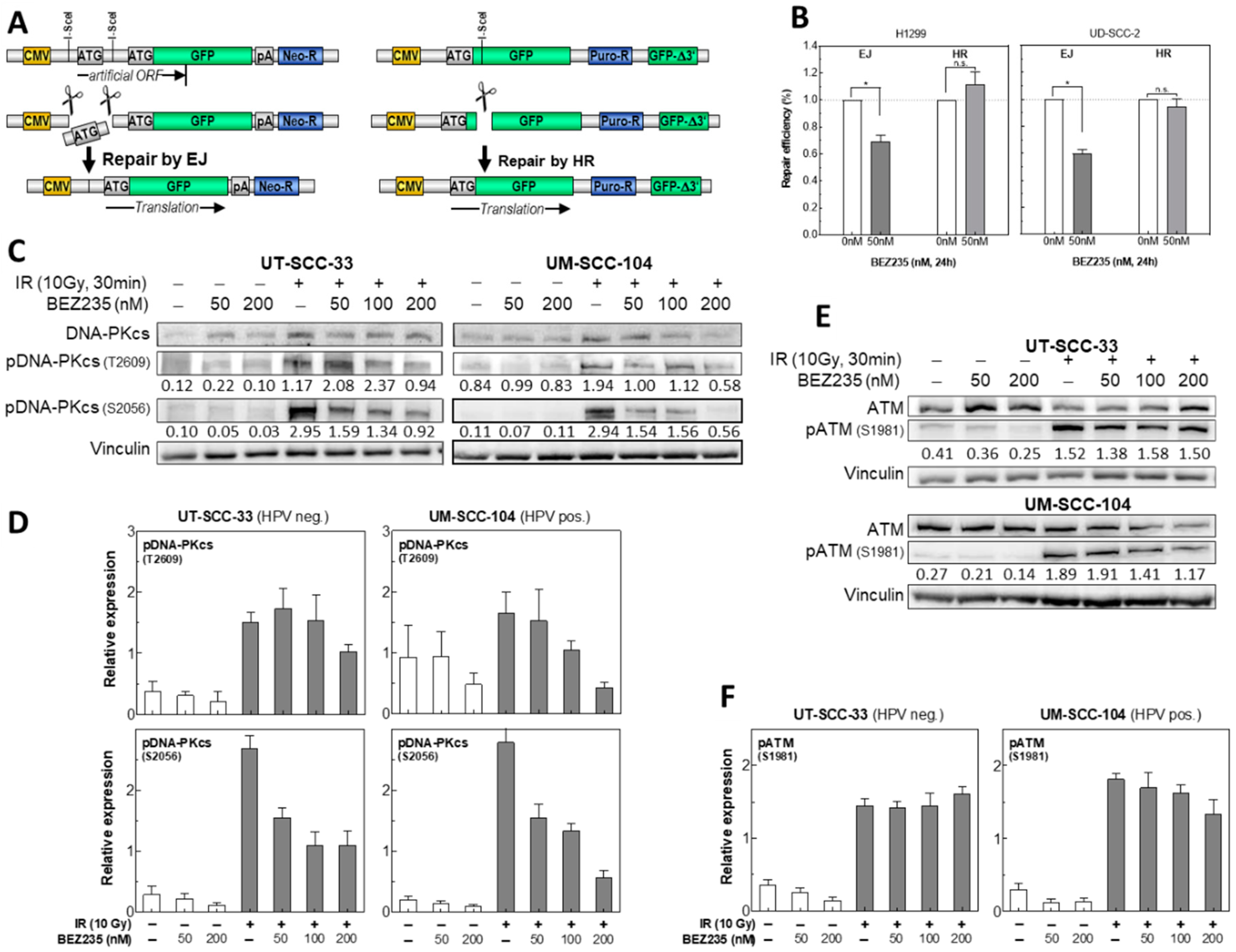
2.5. BEZ235 Suppresses NHEJ Due to an Impaired Phosphorylation of DNA-PKcs at S2056
2.6. BEZ235 Causes Radiosensitization, Which Is Most Pronounced in G0 and G1
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture Conditions
4.2. Radiation and BEZ235 Treatment
4.3. Clonogenic Assay
4.4. Cell Cycle Distribution
4.5. Detection of Apoptosis
4.6. Synchronization of Cells
4.7. Immunofluorescent Microscopy
4.8. Western Blot Analysis
4.9. DSB Reporter Gene Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: Globocan Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. Ca A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cramer, J.D.; Burtness, B.; Le, Q.T.; Ferris, R.L. The Changing Therapeutic Landscape of Head and Neck Cancer. Nat. Rev. Clin. Oncol. 2019, 16, 669–683. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Tan, P.F.; Zhang, Q.; Ang, K.K.; Weber, R.S.; Rosenthal, D.I.; Soulieres, D.; Kim, H.; Silverman, C.; Raben, A.; Galloway, T.J.; et al. Randomized Phase Iii Trial to Test Accelerated Versus Standard Fractionation in Combination with Concurrent Cisplatin for Head and Neck Carcinomas in the Radiation Therapy Oncology Group 0129 Trial: Long-Term Report of Efficacy and Toxicity. J. Clin. Oncol. 2014, 32, 3858–3866. [Google Scholar] [CrossRef] [PubMed]
- A Bonner, J.; Harari, P.M.; Giralt, J.; Cohen, R.B.; Jones, C.U.; Sur, R.K.; Raben, D.; Baselga, J.; A Spencer, S.; Zhu, J.; et al. Radiotherapy Plus Cetuximab for Locoregionally Advanced Head and Neck Cancer: 5-Year Survival Data from a Phase 3 Randomised Trial, and Relation between Cetuximab-Induced Rash and Survival. Lancet Oncol. 2010, 11, 21–28. [Google Scholar] [CrossRef]
- Kriegs, M.; Kasten-Pisula, U.; Riepen, B.; Hoffer, K.; Struve, N.; Myllynen, L.; Braig, F.; Binder, M.; Rieckmann, T.; Grenman, R.; et al. Radiosensitization of Hnscc Cells by Egfr Inhibition Depends on the Induction of Cell Cycle Arrests. Oncotarget 2016, 7, 45122–45133. [Google Scholar] [CrossRef] [Green Version]
- Kriegs, M.; Gurtner, K.; Can, Y.; Brammer, I.; Rieckmann, T.; Oertel, R.; Wysocki, M.; Dorniok, F.; Gal, A.; Grob, T.J.; et al. Radiosensitization of Nsclc Cells by Egfr Inhibition Is the Result of an Enhanced P53-Dependent G1 Arrest. Radiother. Oncol. 2015, 115, 120–127. [Google Scholar] [CrossRef]
- Stransky, N.; Egloff, A.M.; Tward, A.D.; Kostic, A.D.; Cibulskis, K.; Sivachenko, A.; Kryukov, G.V.; Lawrence, M.S.; Sougnez, C.; McKenna, A.; et al. The Mutational Landscape of Head and Neck Squamous Cell Carcinoma. Science 2011, 333, 1157–1160. [Google Scholar] [CrossRef] [Green Version]
- Iglesias-Bartolome, R.; Martin, D.; Gutkind, J.S. Exploiting the Head and Neck Cancer Oncogenome: Widespread Pi3k-Mtor Pathway Alterations and Novel Molecular Targets. Cancer Discov. 2013, 3, 722–725. [Google Scholar] [CrossRef] [Green Version]
- Lui, V.W.; Hedberg, M.L.; Li, H.; Vangara, B.S.; Pendleton, K.; Zeng, Y.; Lu, Y.; Zhang, Q.; Du, Y.; Gilbert, B.R.; et al. Frequent Mutation of the Pi3k Pathway in Head and Neck Cancer Defines Predictive Biomarkers. Cancer Discov. 2013, 3, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas, Network. Comprehensive Genomic Characterization of Head and Neck Squamous Cell Carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef] [Green Version]
- Vander Broek, R.; Mohan, S.; Eytan, D.F.; Chen, Z.; Van Waes, C. The Pi3k/Akt/Mtor Axis in Head and Neck Cancer: Functions, Aberrations, Cross-Talk, and Therapies. Oral. Dis. 2015, 21, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Toulany, M.; Lee, K.-J.; Fattah, K.R.; Lin, Y.-F.; Fehrenbacher, B.; Schaller, M.; Chen, B.P.; Chen, D.J.; Rodemann, H.P. Akt Promotes Post-Irradiation Survival of Human Tumor Cells through Initiation, Progression, and Termination of DNA-Pkcs-Dependent DNA Double-Strand Break Repair. Mol. Cancer Res. 2012, 10, 945–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueck, K.; Rebholz, S.; Harati, M.D.; Rodemann, H.P.; Toulany, M. Akt1 Stimulates Homologous Recombination Repair of DNA Double-Strand Breaks in a Rad51-Dependent Manner. Int. J. Mol. Sci. 2017, 18, 2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeck, S.; Al-Refae, K.; Riffkin, H.; Wiel, G.; Handrick, R.; Klein, D.; Iliakis, G.; Jendrossek, V. Activating Akt1 Mutations Alter DNA Double Strand Break Repair and Radiosensitivity. Sci. Rep. 2017, 7, 42700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maira, S.-M.; Stauffer, F.; Brueggen, J.; Furet, P.; Schnell, C.; Fritsch, C.; Brachmann, S.; Chène, P.; De Pover, A.; Schoemaker, K.; et al. Identification and Characterization of Nvp-Bez235, a New Orally Available Dual Phosphatidylinositol 3-Kinase/Mammalian Target of Rapamycin Inhibitor with Potent in Vivo Antitumor Activity. Mol. Cancer Ther. 2008, 7, 1851–1863. [Google Scholar] [CrossRef] [Green Version]
- Toledo, L.I.; Murga, M.; Zur, R.; Soria, R.; Rodriguez, A.; Martínez, S.; Oyarzabal, J.; Pastor, J.; Bischoff, J.R.; Fernandez-Capetillo, O. A Cell-Based Screen Identifies Atr Inhibitors with Synthetic Lethal Properties for Cancer-Associated Mutations. Nat. Struct. Mol. Biol. 2011, 18, 721–727. [Google Scholar] [CrossRef] [Green Version]
- Konstantinidou, G.; Bey, E.A.; Rabellino, A.; Schuster, K.; Maira, M.S.; Gazdar, A.F.; Amici, A.; Boothman, D.A.; Scaglioni, P.P. Dual Phosphoinositide 3-Kinase/Mammalian Target of Rapamycin Blockade Is an Effective Radiosensitizing Strategy for the Treatment of Non-Small Cell Lung Cancer Harboring K-Ras Mutations. Cancer Res. 2009, 69, 7644–7652. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-H.; Wei, M.-F.; Wang, C.-W.; Lee, H.-W.; Pan, S.-L.; Gao, M.; Kuo, S.-H.; Cheng, A.-L.; Teng, C.-M. Dual Phosphoinositide 3-Kinase/Mammalian Target of Rapamycin Inhibitor Is an Effective Radiosensitizer for Colorectal Cancer. Cancer Lett. 2015, 357, 582–590. [Google Scholar] [CrossRef]
- Mukherjee, B.; Tomimatsu, N.; Amancherla, K.; Camacho, C.V.; Pichamoorthy, N.; Burma, S. The Dual Pi3k/Mtor Inhibitor Nvp-Bez235 Is a Potent Inhibitor of Atm-and DNA-Pkcs-Mediated DNA Damage Responses. Neoplasia 2012, 14, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Del Alcazar, C.R.G.; Hardebeck, M.C.; Mukherjee, B.; Tomimatsu, N.; Gao, X.; Yan, J.; Xie, X.J.; Bachoo, R.; Li, L.; Burma, S.; et al. Inhibition of DNA Double-Strand Break Repair by the Dual Pi3k/Mtor Inhibitor Nvp-Bez235 as a Strategy for Radiosensitization of Glioblastoma. Clin. Cancer Res. 2014, 20, 1235–1248. [Google Scholar] [CrossRef] [Green Version]
- Fokas, E.; Yoshimura, M.; Prevo, R.; Higgins, G.; Hackl, W.; Maira, S.-M.; Bernhard, E.J.; McKenna, W.G.; Muschel, R.J. Nvp-Bez235 and Nvp-Bgt226, Dual Phosphatidylinositol 3-Kinase/Mammalian Target of Rapamycin Inhibitors, Enhance Tumor and Endothelial Cell Radiosensitivity. Radiat. Oncol. 2012, 7, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuger, S.; Graus, D.; Brendtke, R.; Günther, N.; Katzer, A.; Lutyj, P.; Polat, B.; Chatterjee, M.; Sukhorukov, V.L.; Flentje, M.; et al. Radiosensitization of Glioblastoma Cell Lines by the Dual Pi3k and Mtor Inhibitor Nvp-Bez235 Depends on Drug-Irradiation Schedule. Transl. Oncol. 2013, 6, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.-C.; Hung, S.-K.; Lin, H.-Y.; Chiou, W.-Y.; Lee, M.-S.; Liao, H.-F.; Huang, H.-B.; Ho, H.-C.; Su, Y.-C. Targeting the Pi3k/Akt/Mtor Signaling Pathway as an Effectively Radiosensitizing Strategy for Treating Human Oral Squamous Cell Carcinoma in Vitro and in Vivo. Oncotarget 2017, 8, 68641–68653. [Google Scholar] [CrossRef] [PubMed]
- Arenz, A.; Ziemann, F.; Mayer, C.; Wittig, A.; Dreffke, K.; Preising, S.; Wagner, S.; Klussmann, J.-P.; Engenhart-Cabillic, R.; Wittekindt, C. Increased Radiosensitivity of Hpv-Positive Head and Neck Cancer Cell Lines Due to Cell Cycle Dysregulation and Induction of Apoptosis. Strahlenther. Onkol. 2014, 190, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Rieckmann, T.; Tribius, S.; Grob, T.J.; Meyer, F.; Busch, C.-J.; Petersen, C.; Dikomey, E.; Kriegs, M. Hnscc Cell Lines Positive for Hpv and P16 Possess Higher Cellular Radiosensitivity Due to an Impaired Dsb Repair Capacity. Radiother. Oncol. 2013, 107, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Ziemann, F.; Seltzsam, S.; Dreffke, K.; Preising, S.; Arenz, A.; Subtil, F.S.; Rieckmann, T.; Engenhart-Cabillic, R.; Dikomey, E.; Wittig, A. Roscovitine Strongly Enhances the Effect of Olaparib on Radiosensitivity for Hpv Neg. But Not for Hpv Pos. Hnscc Cell Lines. Oncotarget 2017, 8, 105170–105183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessi, D.R.; James, S.R.; Downes, C.P.; Holmes, A.B.; Gaffney, P.R.; Reese, C.B.; Cohen, P. Characterization of a 3-Phosphoinositide-Dependent Protein Kinase Which Phosphorylates and Activates Protein Kinase Balpha. Curr. Biol. 1997, 7, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Sarbassov, D.D.; Guertin, D.A.; Ali, S.M.; Sabatini, D.M. Phosphorylation and Regulation of Akt/Pkb by the Rictor-Mtor Complex. Science 2005, 307, 1098–1101. [Google Scholar] [CrossRef] [Green Version]
- Yung, H.W.; Charnock-Jones, D.S.; Burton, G.J. Regulation of Akt Phosphorylation at Ser473 and Thr308 by Endoplasmic Reticulum Stress Modulates Substrate Specificity in a Severity Dependent Manner. PLoS ONE 2011, 6, e17894. [Google Scholar] [CrossRef]
- Alessi, D.R.; Andjelkovic, M.; Caudwell, B.; Cron, P.; Morrice, N.; Cohen, P.; Hemmings, B.A. Mechanism of Activation of Protein Kinase B by Insulin and Igf-1. EMBO J. 1996, 15, 6541–6551. [Google Scholar] [CrossRef] [Green Version]
- Freudlsperger, C.; Horn, D.; Weißfuß, S.; Weichert, W.; Weber, K.J.; Saure, D.; Sharma, S.; Dyckhoff, G.; Grabe, N.; Hoffmann, J.; et al. Phosphorylation of Akt(Ser473) Serves as an Independent Prognostic Marker for Radiosensitivity in Advanced Head and Neck Squamous Cell Carcinoma. Int. J. Cancer 2015, 136, 2775–2785. [Google Scholar] [CrossRef] [PubMed]
- Eke, I.; Makinde, A.Y.; Aryankalayil, M.J.; Sandfort, V.; Palayoor, S.T.; Rath, B.H.; Liotta, L.; Pierbon, M.; Petricoin, E.F.; Stommel, J.M.; et al. Exploiting Radiation-Induced Signaling to Increase the Susceptibility of Resistant Cancer Cells to Targeted Drugs: Akt and Mtor Inhibitors as an Example. Mol. Cancer 2018, 17, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Slingerland, J.M. Multiple Roles of the Pi3k/Pkb (Akt) Pathway in Cell Cycle Progression. Cell Cycle 2003, 2, 339–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köcher, S.; Rieckmann, T.; Rohaly, G.; Mansour, W.Y.; Dikomey, E.; Dornreiter, I.; Dahm-Daphi, J. Radiation-Induced Double-Strand Breaks Require Atm but Not Artemis for Homologous Recombination During S-Phase. Nucleic Acids Res. 2012, 40, 8336–8347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieckmann, T.; Kriegs, M.; Nitsch, L.; Hoffer, K.; Rohaly, G.; Kocher, S.; Petersen, C.; Dikomey, E.; Dornreiter, I.; Dahm-Daphi, J. P53 Modulates Homologous Recombination at I-Scei-Induced Double-Strand Breaks through Cell-Cycle Regulation. Oncogene 2013, 32, 968–975. [Google Scholar] [CrossRef] [Green Version]
- Mansour, W.Y.; Schumacher, S.; Rosskopf, R.; Rhein, T.; Schmidt-Petersen, F.; Gatzemeier, F.; Haag, F.; Borgmann, K.; Willers, H.; Dahm-Daphi, J. Hierarchy of Nonhomologous End-Joining, Single-Strand Annealing and Gene Conversion at Site-Directed DNA Double-Strand Breaks. Nucleic Acids Res. 2008, 36, 4088–4098. [Google Scholar] [CrossRef] [Green Version]
- Toulany, M.; Maier, J.; Iida, M.; Rebholz, S.; Holler, M.; Grottke, A.; Jüker, M.; Wheeler, D.L.; Rothbauer, U.; Rodemann, H.P. Akt1 and Akt3 but Not Akt2 through Interaction with DNA-Pkcs Stimulate Proliferation and Post-Irradiation Cell Survival of K-Ras-Mutated Cancer Cells. Cell Death Discov. 2017, 3, 17072. [Google Scholar] [CrossRef]
- Bakr, A.; Oing, C.; Köcher, S.; Borgmann, K.; Dornreiter, I.; Petersen, C.; Dikomey, E.; Mansour, W. Involvement of Atm in Homologous Recombination after End Resection and Rad51 Nucleofilament Formation. Nucleic Acids Res. 2015, 43, 3154–3166. [Google Scholar] [CrossRef] [Green Version]
- Carlo, M.I.; Molina, A.M.; Lakhman, Y.; Patil, S.; Woo, K.; DeLuca, J.; Lee, C.-H.; Hsieh, J.J.; Feldman, D.R.; Motzer, R.J.; et al. A Phase Ib Study of Bez235, a Dual Inhibitor of Phosphatidylinositol 3-Kinase (Pi3k) and Mammalian Target of Rapamycin (Mtor), in Patients with Advanced Renal Cell Carcinoma. Oncologist 2016, 21, 787–788. [Google Scholar] [CrossRef] [Green Version]
- Salazar, R.; Garcia-Carbonero, R.; Libutti, S.K.; Hendifar, A.E.; Custodio, A.; Guimbaud, R.; Lombard-Bohas, C.; Ricci, S.; Klümpen, H.-J.; Reed, N. Phase Ii Study of Bez235 Versus Everolimus in Patients with Mammalian Target of Rapamycin Inhibitor-Naive Advanced Pancreatic Neuroendocrine Tumors. Oncologist 2018, 23, 766. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-H.; Wang, C.-W.; Wei, M.-F.; Tzeng, Y.-S.; Lan, K.-H.; Cheng, A.-L.; Kuo, S.-H. Maintenance Bez235 Treatment Prolongs the Therapeutic Effect of the Combination of Bez235 and Radiotherapy for Colorectal Cancer. Cancers 2019, 11, 1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuger, S.; Cörek, E.; Polat, B.; Kämmerer, U.; Flentje, M.; Djuzenova, C.S. Novel Pi3k and Mtor Inhibitor Nvp-Bez235 Radiosensitizes Breast Cancer Cell Lines under Normoxic and Hypoxic Conditions. Breast Cancer 2014, 8, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziemann, F.; Arenz, A.; Preising, S.; Wittekindt, C.; Klussmann, J.P.; Engenhart-Cabillic, R.; Wittig, A. Increased Sensitivity of Hpv-Positive Head and Neck Cancer Cell Lines to X-Irradiation +/-Cisplatin Due to Decreased Expression of E6 and E7 Oncoproteins and Enhanced Apoptosis. Am. J. Cancer Res. 2015, 5, 1017–1031. [Google Scholar] [PubMed]
- Seltzsam, S.; Ziemann, F.; Dreffke, K.; Preising, S.; Arenz, A.; Schötz, U.; Engenhart-Cabillic, R.; Dikomey, E.; Wittig, A. In Hpv-Positive Hnscc Cells, Functional Restoration of the P53/P21 Pathway by Proteasome Inhibitor Bortezomib Does Not Affect Radio-or Chemosensitivity. Transl. Oncol. 2019, 12, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Cerniglia, G.J.; Karar, J.; Tyagi, S.; Christofidou-Solomidou, M.; Rengan, R.; Koumenis, C.; Maity, A. Inhibition of Autophagy as a Strategy to Augment Radiosensitization by the Dual Phosphatidylinositol 3-Kinase/Mammalian Target of Rapamycin Inhibitor Nvp-Bez235. Mol. Pharm. 2012, 82, 1230–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fokas, E.; Im, J.H.; Hill, S.; Yameen, S.; Stratford, M.; Beech, J.; Hackl, W.; Michel Maira, S.; Bernhard, E.J.; Muschel, R.J.; et al. Dual Inhibition of the Pi3k/Mtor Pathway Increases Tumor Radiosensitivity by Normalizing Tumor Vasculature. Cancer Res. 2012, 72, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Helleday, T.; Lo, J.; Van Gent, D.C.; Engelward, B.P. DNA Double-Strand Break Repair: From Mechanistic Understanding to Cancer Treatment. DNA Repair 2007, 6, 923–935. [Google Scholar] [CrossRef]
- Meek, K.; Douglas, P.; Cui, X.; Ding, Q.; Lees-Miller, S.P. Trans Autophosphorylation at DNA-Dependent Protein Kinase’s Two Major Autophosphorylation Site Clusters Facilitates End Processing but Not End Joining. Mol. Cell. Biol. 2007, 27, 3881–3890. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.P.; Uematsu, N.; Kobayashi, J.; Lerenthal, Y.; Krempler, A.; Yajima, H.; Löbric, M.; Shiloh, Y.; Chen, D.J. Ataxia Telangiectasia Mutated (Atm) Is Essential for DNA-Pkcs Phosphorylations at the Thr-2609 Cluster Upon DNA Double Strand Break. J. Biol. Chem. 2007, 282, 6582–6587. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.-J.; Long, L.-M.; Yang, N.; Zhang, Q.-Q.; Ji, W.-J.; Zhao, J.-H.; Qin, Z.-H.; Wang, Z.; Chen, G.; Liang, Z.-Q. Nvp-Bez235, a Novel Dual Pi3k/Mtor Inhibitor, Enhances the Radiosensitivity of Human Glioma Stem Cells in Vitro. Acta Pharmacol. Sin. 2013, 34, 681–690. [Google Scholar] [CrossRef] [Green Version]
- Dirks, W.G.; MacLeod, R.A.F.; Nakamura, Y.; Kohara, A.; Reid, Y.; Milch, H.; Drexler, H.G.; Mizusawa, H. Cell Line Cross-Contamination Initiative: An Interactive Reference Database of Str Profiles Covering Common Cancer Cell Lines. Int. J. Cancer 2010, 126, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Uphoff, C.C.; Drexler, H.G. Comparative Pcr Analysis for Detection of Mycoplasma Infections in Continuous Cell Lines. In Vitro Cell. Dev. Biol. Anim. 2002, 38, 79–85. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schötz, U.; Balzer, V.; Brandt, F.-W.; Ziemann, F.; Subtil, F.S.B.; Rieckmann, T.; Köcher, S.; Engenhart-Cabillic, R.; Dikomey, E.; Wittig, A.; et al. Dual PI3K/mTOR Inhibitor NVP-BEZ235 Enhances Radiosensitivity of Head and Neck Squamous Cell Carcinoma (HNSCC) Cell Lines Due to Suppressed Double-Strand Break (DSB) Repair by Non-Homologous End Joining. Cancers 2020, 12, 467. https://doi.org/10.3390/cancers12020467
Schötz U, Balzer V, Brandt F-W, Ziemann F, Subtil FSB, Rieckmann T, Köcher S, Engenhart-Cabillic R, Dikomey E, Wittig A, et al. Dual PI3K/mTOR Inhibitor NVP-BEZ235 Enhances Radiosensitivity of Head and Neck Squamous Cell Carcinoma (HNSCC) Cell Lines Due to Suppressed Double-Strand Break (DSB) Repair by Non-Homologous End Joining. Cancers. 2020; 12(2):467. https://doi.org/10.3390/cancers12020467
Chicago/Turabian StyleSchötz, Ulrike, Viola Balzer, Friedrich-Wilhelm Brandt, Frank Ziemann, Florentine S.B. Subtil, Thorsten Rieckmann, Sabrina Köcher, Rita Engenhart-Cabillic, Ekkehard Dikomey, Andrea Wittig, and et al. 2020. "Dual PI3K/mTOR Inhibitor NVP-BEZ235 Enhances Radiosensitivity of Head and Neck Squamous Cell Carcinoma (HNSCC) Cell Lines Due to Suppressed Double-Strand Break (DSB) Repair by Non-Homologous End Joining" Cancers 12, no. 2: 467. https://doi.org/10.3390/cancers12020467