 Tatiana Rodrigues Fraga
Tatiana Rodrigues Fraga Lourdes Isaac
Lourdes Isaac Angela Silva Barbosa
Angela Silva Barbosa- 1Laboratory of Complement, Department of Immunology, Institute of Biomedical Sciences, University of São Paulo, São Paulo, Brazil
- 2Laboratory of Bacteriology, Instituto Butantan, São Paulo, Brazil
Leptospirosis is a neglected infectious disease caused by spirochetes from the genus Leptospira. Pathogenic microorganisms, notably those which reach the blood circulation such as Leptospira, have evolved multiple strategies to escape the host complement system, which is important for innate and acquired immunity. Leptospira avoid complement-mediated killing through: (i) recruitment of host complement regulators; (ii) acquisition of host proteases that cleave complement proteins on the bacterial surface; and, (iii) secretion of proteases that inactivate complement proteins in the Leptospira surroundings. The recruitment of host soluble complement regulatory proteins includes the acquisition of Factor H (FH) and FH-like-1 (alternative pathway), C4b-binding protein (C4BP) (classical and lectin pathways), and vitronectin (Vn) (terminal pathway). Once bound to the leptospiral surface, FH and C4BP retain cofactor activity of Factor I in the cleavage of C3b and C4b, respectively. Vn acquisition by leptospires may result in terminal pathway inhibition by blocking C9 polymerization. The second evasion mechanism lies in plasminogen (PLG) binding to the leptospiral surface. In the presence of host activators, PLG is converted to enzymatically active plasmin, which is able to degrade C3b, C4b, and C5 at the surface of the pathogen. A third strategy used by leptospires to escape from complement system is the active secretion of proteases. Pathogenic, but not saprophytic leptospires, are able to secrete metalloproteases that cleave C3 (central complement molecule), Factor B (alternative pathway), and C4 and C2 (classical and lectin pathways). The purpose of this review is to fully explore these complement evasion mechanisms, which act together to favor Leptospira survival and multiplication in the host.
Introduction
Spirochetes of the genus Leptospira are highly motile Gram-negative bacteria that cause a worldwide zoonosis (1). This bacterium colonizes a wide range of hosts including humans, domestic and wild animal species. Patients with leptospirosis may present either very mild symptoms or subclinical disease (80–90% of infections) or a more severe illness characterized by jaundice, acute renal failure and bleeding (Weil’s disease), or pulmonary hemorrhage syndrome [reviewed in Ref. (2)].
The genus Leptospira comprises bacteria having distinct ecological adaptations: exclusively non-pathogenic free-living species, exclusively host-dependent organisms and pathogenic species capable of surviving both inside and outside the host for long periods (3). Molecular phylogenetic analysis of 16S rRNA gene sequences allowed clustering of Leptospira species into three groups, comprising pathogens, non-pathogens, and an intermediate group (4).
Upon infection, pathogenic leptospires spread and propagate in susceptible hosts because they have evolved diverse immune evasion strategies. Conversely, saprophytic Leptospira strains are highly susceptible to serum bactericidal activity, an observation already made by Johnson and Muschel in the mid-1960s (5). Since our insights into complement evasion mechanisms of Leptospira have substantially increased during the last 10 years, we aim here to provide a comprehensive overview of the interactions of this relevant human pathogen with the complement system.
The Role of the Complement System in the Host’s Defense Against Pathogens
The complement system is composed of more than 50 plasma proteins and receptors. Traditionally considered as one of the first lines of defense against invading microorganisms due to its opsonic, inflammatory, and lytic activities, complement roles extend far beyond pathogen killing [reviewed in Ref. (6)]. Complement effector functions result from activation of three different pathways: classical, alternative, and/or lectin pathways (CP, AP, and LP, respectively). While the AP and LP participate in the innate immunity, the CP is generally activated by the presence of IgG or IgM specifically bound to antigens. The AP is initiated by the spontaneous hydrolysis of an intra-chain thioester bond located in the C3 molecule, while the LP is activated when lectins, such as mannose-binding lectin or ficolins, bind to carbohydrates commonly found on microorganisms’ surfaces. During activation, fragments C3b and C4b are generated and they bind covalently to acceptor surfaces such as immune complexes, foreign, and host cells located on the vicinity of the activation site. On these surfaces, C3 and C5 convertases are formed which further lead to the formation of the membrane attack complex culminating with microorganism lysis. As a consequence of activation, particles opsonized with iC3b, C3b, and C4b are more efficiently internalized by neutrophils, monocytes, and macrophages once bound to complement receptors present on these cells’ membranes. CR2 promotes activation and proliferation of B lymphocytes in the presence of C3d/C3dg fragments covalently bound to antigens inducing the production of antibodies. In addition, C3a and C5a fragments are important anaphylatoxins. They are also chemoattractant factors for inflammatory cells [reviewed in Ref. (7)]. In order to protect the host against self-damage, complement activation is tightly controlled at all stages of the cascade by several soluble and cell surface regulators. C1 inhibitor, Factor I (FI), Factor H (FH), and C4b-binding protein (C4BP) are soluble complement regulators whereas complement receptor type 1 (CR1 or CD35), membrane cofactor protein (MCP or CD46), decay accelerator factor (DAF or CD55), and CD59 are cell-anchored regulatory receptors [reviewed in Ref. (7, 8)].
Complement Evasion Strategies by Leptospira
Pathogens use a range of strategies to avoid complement attack, and Leptospira is no exception to this phenomenon. While pathogenic Leptospira strains resist complement-mediated killing, saprophyte Leptospira strains are highly susceptible to serum killing (9, 10). Concerning the group of leptospires of intermediate pathogenicity, such as Leptospira licerasiae, nothing is known about their response to complement. Pathogenic Leptospira escape from complement-mediated killing through: (i) recruitment of host complement regulators; (ii) acquisition of host proteases that cleave complement proteins on the bacterial surface; and (iii) secretion of proteases that inactivate complement in the Leptospira surroundings (Figure 1). These mechanisms are universal strategies employed by diverse pathogens including bacteria, viruses, and fungi to circumvent complement attack [reviewed in Ref. (11)].
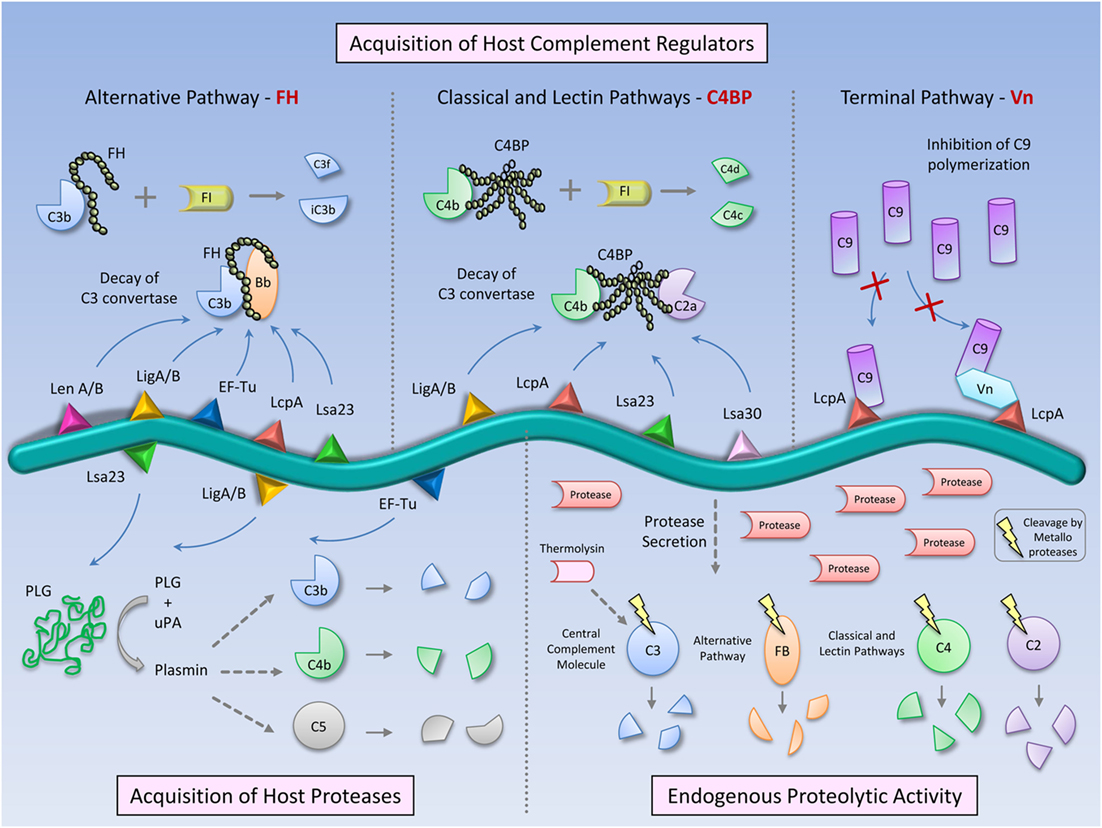
Figure 1. Complement evasion strategies of pathogenic Leptospira. To circumvent the complement system, Leptospira has evolved different immune evasion strategies: (i) acquisition of host soluble complement regulators: Factor H (FH)—AP regulator, C4b-binding protein (C4BP) —CP and LP regulator, and vitronectin (Vn) —terminal pathway regulator. FH and C4BP accelerate the decay of the C3 convertases (C3bBb and C4b2a, respectively) and act as cofactors for Factor I in C3b and C4b cleavages. Vn and the leptospiral protein LcpA bind C9 and inhibit its polymerization, thus potentially blocking MAC formation; (ii) acquisition of host proteases: pathogenic Leptospira binds plasminogen, which in the presence of activators, such as Urokinase-type plasminogen activator (uPA), is converted in the enzymatically active plasmin. This serine protease cleaves C3b, C4b, and C5, promoting a downregulation of complement activation on the Leptospira surface, and (iii) Direct inactivation of complement proteins by Leptospira endogenous proteases. Metalloproteases secreted by pathogenic Leptospira strains are able to cleave and inactivate the complement proteins: C3 (central complement molecule), Factor B (from AP), and C2 and C4 (CP and LP). Thermolysin is one of the proteases responsible for these cleavages, degrading C3. The combination of host-derived and endogenous factors from pathogenic Leptospira enables the bacteria to successfully establish infection and colonize target organs of the host.
Recruitment of Mammalian Host Complement Soluble Regulatory Proteins
Pathogenic Leptospira are potentially able to control all pathways of the complement system by acquiring soluble negative host regulators. Control of the AP is achieved by interaction of Leptospira with FH, a 155 kDa plasma glycoprotein [443 ± 106 μg/mL in human serum (12)] composed of 20 globular domains (termed complement control protein domains, CCPs). FH inhibits AP activation by preventing binding of Factor B (FB) to C3b, by accelerating the decay of the C3-convertase C3bBb and by acting as a cofactor for the cleavage of C3b by FI (13–15). Serum-resistant Leptospira strains bind three members of the FH family: FH itself, Factor H-like protein 1 (FHL-1), and Factor H-related protein 1 (FHR-1) (9, 16). Once bound to the Leptospira surface, FH remains functional and promotes FI-mediated cleavage of C3b, thus generating the iC3b fragment (9, 16). Moreover, Leptospira survival in FH-depleted serum was shown to be impaired by 60%, and reconstitution of this serum with purified FH up to physiological concentrations restored bacterial survival in a dose-dependent manner, further supporting a role for FH in Leptospira serum resistance (16). Control of the CP by pathogenic Leptospira is mediated by surface-bound C4BP, a 570 kDa plasma glycoprotein [335 ± 83 μg/ml (12)] that is found in three isoforms with different subunit composition. The major isoform, α7/β1, is a complex of seven α-chains and one β-chain. The other C4BP isoforms in plasma are α7/β0 and α6/β1. Each α-chain is comprised of eight CCPs, and the β-chain is comprised of three CCPs (17). C4BP inhibits CP and LP activation by interfering with the assembly and decay of the C3 convertase (C4bC2a) and by acting as a cofactor for FI in the proteolytic inactivation of C4b (18, 19). Both virulent and culture-attenuated Leptospira strains express ligands for C4BP, in contrast to non-pathogenic strains, which have been shown to bind insignificant amounts of this complement regulator (10). As expected, C4b is more efficiently cleaved by FI in the presence of C4BP bound to Leptospira interrogans virulent strains, which may probably explain their higher survival rate in normal human serum as compared to culture-attenuated strains (10). Leptospires also acquire vitronectin (Vn) on their surfaces (20). Vn is a glycoprotein that circulates in the bloodstream as a monomer [65–75 kDa, 104 ± 25 μg/mL (12)] or is deposited in the extracellular matrix (ECM) as a multimer that interacts with several macromolecular components, including glycosaminoglycans and collagens (21, 22). Vn plays multiple roles in many biological processes including the regulation of the terminal pathway of complement by inhibiting C5b7 complex formation and C9 polymerization. Once bound to the bacterial surface, it may protect the microorganism against lysis by impairing MAC formation. A number of strains belonging to different Leptospira species have been shown to interact with human Vn (20). Acquisition of this terminal pathway regulatory protein may assist Leptospira to evade complement attack.
Leptospira Ligands for Host Complement Regulators
Pathogenic Leptospira bind soluble host complement regulators via surface proteins and multiple ligands for those regulators have been described. The most extensively characterized complement evasion molecules from Leptospira are (i) leptospiral endostatin-like proteins A and B [LenA and LenB (23, 24)], (ii) Leptospira immunoglobulin-like (Lig) proteins A and B [LigA and LigB (16, 25)], and (iii) Leptospiral complement regulator-acquiring protein A [LcpA (26)]. All of these proteins have been shown to bind more than one complement regulator and seem to be involved not only in immune evasion but also in adhesion and invasion by interacting with ECM and plasma proteins such as plasminogen (PLG) (27).
Leptospira interrogans strains encode up to six distinct paralogous proteins called LenA–F, harboring domains that presumably share structural and functional similarities with mammalian endostatins (24). Two proteins of this family, LenA (formerly called LfhA and Lsa24) and LenB, have affinities for complement regulators. While LenA binds both FH and FHR-1, LenB has been shown to interact only with FH (23, 24).
LigA and LigB are multifunctional proteins capable of interacting with the ECM, cell lines, and complement regulators in vitro. The family of Lig proteins is composed of LigA, LigB, and LigC, which respectively consist of 13, 12, and 13 Ig-like domains. In certain Leptospira species ligC is a pseudogene (28). The lig genes are present only in pathogenic Leptospira species, and they are expressed during mammalian infection (28). Recombinant LigA and LigB bind FH, FHL-1, FHR-1, and C4BP, thus potentially allowing control of all complement activation pathways (16). FH CCP5 and CCP20 domains interact with both LigA and LigB (16). C4BP CCP4, CCP7, and CCP8 domains are involved in the interaction with both LigA and LigB (29). Fine mapping of the LigA and LigB domains involved in binding to C4BP has demonstrated that interactions occur through the bacterial immunoglobulin-like (Big) domains 7 and 8 (LigA7–8 and LigB7–8) of both LigA and LigB and also through LigB9–10 (29). As FH and C4BP do not compete for binding to Lig proteins, they probably have distinct binding sites on these molecules and may then interact with their targets simultaneously (16). It has been shown that ectopic LigB expression promotes survival of the saprophyte Leptospira biflexa in normal human serum (30). LigB binds C3b and C4b directly through repeats 9–11 (LigB9–11) and inhibits both the alternative and classical pathways in hemolytic assays with erythrocytes (30). Given the susceptibility of non-pathogenic Leptospira to the alternative pathway (9, 10), the increased resistance of ligB-transformed L. biflexa to complement killing may be attributed to the acquisition of C3b and FH by these bacteria (30). Further studies extended this observation by demonstrating that expression of both ligA and ligB genes enhances L. biflexa’s resistance to serum killing, as demonstrated by a reduced MAC deposition on lig-transformed L. biflexa compared to the wild type strain (31).
Pathogenic Leptospira species also bind host’s negative complement regulators through a 20-kDa surface-exposed lipoprotein named LcpA. First described as a C4BP-interacting protein (26), LcpA was later shown to bind FH and Vn as well as the terminal pathway component C9 (20). Usually, microorganisms bind FH via a common site located inside CCP20 (32). LcpA is no exception to this rule, since a monoclonal antibody directed against CCP20 inhibited binding of FH to LcpA (20). CCP7 and CCP8 domains mediate the interaction of C4BP with LcpA (29). Both FH and C4BP have been shown to remain functional once bound to LcpA, thus being able to act as cofactors for FI (20, 26). LcpA also interferes with the terminal pathway of complement by binding to C9, a molecule that has a key role in MAC formation on bacterial cells. In the presence of LcpA, Zn2+-induced C9 polymerization is inhibited in vitro and MAC formation on sheep erythrocytes is partially impaired, preventing cell lysis (20). Competitive binding assays indicate that LcpA interacts with C4BP, FH, and Vn through distinct sites (20).
Based on binding affinities, other Leptospira proteins have been shown to acquire complement regulators (Table 1). Lsa30 binds C4BP whereas Lsa23 binds both C4BP and FH (33, 34). Interestingly, the moonlighting protein EF-Tu, shown to be surface-exposed in Leptospira, also acquires the complement regulator FH (35).
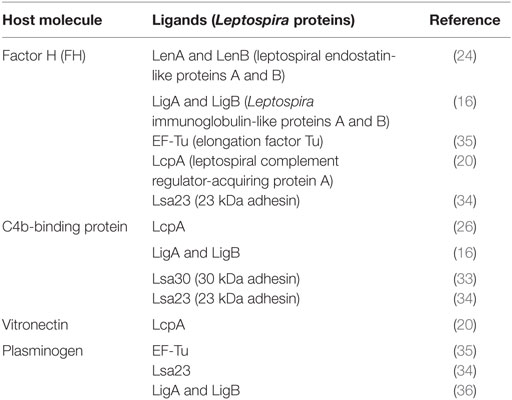
Table 1. Host molecules that interact with Leptospira ligands to evade the complement system.
Acquisition of Host Proteases That Cleave Complement Proteins on the Leptospira Surface
Proteolytic activity is a fundamental tool employed by diverse pathogens to both overcome tissue barriers and evade the immune system (37). Degradation of ECM components favors pathogen spreading and dissemination, while cleavage and inactivation of immune effector molecules dampen the host defense system, allowing an effective establishment of the infection (38, 39).
Pathogenic leptospires circumvent complement attack by the cleavage and inactivation of key complement molecules from the three activation pathways. The degradation of complement proteins may occur indirectly, using host-acquired proteases such as PLG, or directly, by the activity of endogenous proteases produced by pathogenic Leptospira strains, as discussed in the next section.
It is well-known that leptospires are able to bind human PLG (40, 41). PLG is a single-chain glycoprotein (92 kDa) that is a key component of the host fibrinolytic system. This proenzyme is found in plasma and extracellular fluids at concentrations of 180–200 µg/mL (42).
Although both saprophytic and pathogenic leptospires bind purified PLG, only pathogenic strains are able to acquire PLG from human plasma (40, 41). The interaction of PLG with Leptospira is mediated by bacterial membrane proteins (Table 1), and involves lysine residues, which are probably positioned at the PLG kringle domains. Another interesting finding is that Leptospira cell integrity is preserved, since cellular growth is not impaired by PLG binding (40). Once bound to the Leptospira surface and in the presence of urokinase-type plasminogen activator (uPA), PLG is converted to enzymatically active plasmin (40, 41). Plasmin is a serine protease that cleaves diverse important biological substrates, including ECM proteins, like fibrinogen, and complement molecules, such as C3b and C5 (43). In this way, pathogenic leptospires coated with plasmin showed reduced deposition of C3b and IgG on their surface, which was probably related to proteolytic degradation of these molecules, potentially reducing opsonization (44). Furthermore, L. interrogans serovar Pomona also displayed enhanced survival in human serum when bound to plasmin (44), which reinforces its role in complement resistance.
Several Leptospira membrane proteins have been described as PLG ligands (33, 41, 45–56). However, only a few of them were indeed shown to directly interfere with complement activation: the elongation factor Tu [EF-Tu (35)]; LigA and LigB (36) and Leptospira 23 kDa surface adhesion [Lsa23 (34)], whose interactions with PLG resulted in the cleavage of C3b, C4b, and/or C5 (Table 1).
Secretion of Leptospiral Proteases That Directly Inactivate Complement
The Leptospira evasion strategies described until know focused on host molecules hijacked by the pathogen to inactivate complement on its surface. Recently, we demonstrated that pathogenic Leptospira can produce molecules that are able to directly interfere with the complement system, in a manner independent of the host machinery. It was observed that the culture supernatants of pathogenic, but not of saprophytic Leptospira strains, were able to specifically inhibit all the three activation pathways of complement. The inhibitory effect observed could be directly correlated to the proteolytic activity present in these culture supernatants. Indeed, the leptospiral proteases were able to target a wide range of substrates including C3, a key factor in the amplification of the complement cascade, FB from AP, C2, and C4, from CP and LP. These cleavages were observed both with purified complement proteins or normal human serum, which indicates that the leptospiral proteases exert their function in a physiological context and may contribute to bacterial virulence (57).
The proteolytic activity found exclusively in pathogenic Leptospira supernatants was almost completely abolished by 1.10-phenanthroline, indicating a major role of metalloproteases in the degradation of complement proteins. A recombinant metalloprotease from the thermolysin family, present only in Leptospira pathogenic species, seems to contribute to these cleavages, since it was able to degrade the central complement protein C3 (57).
The degradation and functional inactivation of complement is a key strategy for attenuating diverse immune responses that are dependent on the proper activation of this system (58). The secretion of proteases that directly cleave complement proteins may contribute to Leptospira immune evasion, as demonstrated for a wide range of other pathogens (59).
Concluding Remarks
Complement is a precisely regulated system composed of numerous specific factors that are activated in a cascade-like manner. This multifactorial cascade nature provides diverse targets for possible interferences by pathogen-derived evasion molecules (58). Most successful human pathogens have developed multiple parallel mechanisms of evading the complement system (60). Leptospira, which is a highly invasive spirochete, is a good example of a pathogen that employs diverse strategies to circumvent complement activation (Figure 1). The combination of host-derived and endogenous factors enables these spirochetes to successfully establish the infection and colonize target organs of the host. Therefore, Leptospira ligands of host regulators and secreted proteases constitute potential sites for immune interference, either as vaccine candidates or as targets for therapeutic agents in the development of new treatments and prophylactic approaches in leptospirosis.
Author Contributions
TF participated in the drafting of the article and prepared the tables and figures. LI performed a critical revision and approved the final version to be published. AB participated in the drafting of the article and performed a critical revision of the final version.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors cordially thank professor Shaker Chuck Farah for a critical reading of the manuscript.
Funding
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), grants # 2014/00926-3 and 2010/50043-0.
References
1. Bharti AR, Nally JE, Ricaldi JN, Matthias MA, Diaz MM, Lovett MA, et al. Peru-United States Leptospirosis Consortium. Leptospirosis: a zoonotic disease of global importance. Lancet Infect Dis (2003) 3(12):757–71. doi:10.1016/S1473-3099(03)00830-2
2. Fraga TR, Barbosa AS, Isaac L. Leptospirosis: aspects of innate immunity, immunopathogenesis and immune evasion from the complement system. Scand J Immunol (2011) 73(5):408–19. doi:10.1111/j.1365-3083.2010.02505.x
3. Faine S, Adler B, Bolin C, Perolat P. Leptospira and Leptospirosis. Melbourne: MedScience (1999).
4. Levett PN. Systematics of leptospiraceae. In: Adler B, editor. Leptospira and Leptospirosis. Current Topics in Microbiology and Immunology. New York: Springer-Verlag Berlin Heidelberg (2015). p. 11–20.
6. Reis ES, Mastellos DC, Yancopoulou D, Risitano AM, Ricklin D, Lambris JD. Applying complement therapeutics to rare diseases. Clin Immunol (2015) 161(2):225–40. doi:10.1016/j.clim.2015.08.009
7. Ricklin D, Hajishengallis G, Yang K, Lambris JD. Complement: a key system for immune surveillance and homeostasis. Nat Immunol (2010) 11(9):785–97. doi:10.1038/ni.1923
8. Kim DD, Song WC. Membrane complement regulatory proteins. Clin Immunol (2006) 118(2–3):127–36. doi:10.1016/j.clim.2005.10.014
9. Meri T, Murgia R, Stefanel P, Meri S, Cinco M. Regulation of complement activation at the C3-level by serum resistant leptospires. Microb Pathog (2005) 39(4):139–47. doi:10.1016/j.micpath.2005.07.003
10. Barbosa AS, Abreu PA, Vasconcellos SA, Morais ZM, Gonçales AP, Silva AS, et al. Immune evasion of Leptospira species by acquisition of human complement regulator C4BP. Infect Immun (2009) 77(3):1137–43. doi:10.1128/IAI.01310-08
11. Zipfel PF, Hallström T, Riesbeck K. Human complement control and complemente evasion by pathogenic microbes – tipping the balance. Mol Immunol (2013) 56(3):152–60. doi:10.1016/j.molimm.2013.05.222
12. de Paula PF, Barbosa JE, Junior PR, Ferriani VP, Latorre MR, Nudelman V, et al. Ontogeny of complement regulatory proteins – concentrations of factor h, factor I, c4b-binding protein, properdin and vitronectin in healthy children of different ages and in adults. Scand J Immunol (2003) 58(5):572–7. doi:10.1046/j.1365-3083.2003.01326.x
13. Weiler JM, Daha MR, Austen KF, Fearon DT. Control of the amplification convertase of complement by the plasma protein beta1H. Proc Natl Acad Sci U S A (1976) 73(9):3268–72. doi:10.1073/pnas.73.9.3268
14. Whaley K, Ruddy S. Modulation of the alternative complement pathways by beta 1 H globulin. J Exp Med (1976) 144(5):1147–63. doi:10.1084/jem.144.5.1147
15. Pangburn MK, Schreiber RD, Müller-Eberhard HJ. Human complement C3b inactivator: isolation, characterization, and demonstration of an absolute requirement for the serum protein beta1H for cleavage of C3b and C4b in solution. J Exp Med (1977) 146(1):257–70. doi:10.1084/jem.146.1.257
16. Castiblanco-Valencia MM, Fraga TR, Silva LB, Monaris D, Abreu PA, Strobel S, et al. Leptospiral immunoglobulin-like proteins interact with human complement regulators factor H, FHL-1, FHR-1, and C4BP. J Infect Dis (2012) 205(6):995–1004. doi:10.1093/infdis/jir875
17. Blom AM. Structural and functional studies of complement inhibitor C4b-binding protein. Biochem Soc Trans (2002) 30(Pt 6):978–82. doi:10.1042/bst0300978
18. Gigli I, Fujita T, Nussenzweig V. Modulation of the classical pathway C3 convertase by plasma proteins C4 binding protein and C3b inactivator. Proc Natl Acad Sci U S A (1979) 76(12):6596–600. doi:10.1073/pnas.76.12.6596
19. Scharfstein J, Ferreira A, Gigli I, Nussenzweig V. Human C4-binding protein. I. Isolation and characterization. J Exp Med (1978) 148(1):207–22. doi:10.1084/jem.148.1.207
20. da Silva LB, Miragaia LS, Breda LC, Abe CM, Schmidt MC, Moro AM, et al. Pathogenic Leptospira species acquire factor H and vitronectin via the surface protein LcpA. Infect Immun (2015) 83(3):888–97. doi:10.1128/IAI.02844-14
21. Preissner KT, Seiffert D. Role of vitronectin and its receptors in haemostasis and vascular remodeling. Thromb Res (1998) 89(1):1–21. doi:10.1016/S0049-3848(97)00298-3
22. Stockmann A, Hess S, Declerck P, Timpl R, Preissner KT. Multimeric vitronectin. Identification and characterization of conformation-dependent self-association of the adhesive protein. J Biol Chem (1993) 268(30):22874–82.
23. Verma A, Hellwage J, Artiushin S, Zipfel PF, Kraiczy P, Timoney JF, et al. LfhA, a novel factor H-binding protein of Leptospira interrogans. Infect Immun (2006) 74(5):2659–66. doi:10.1128/IAI.74.5.2659-2666.2006
24. Stevenson B, Choy HA, Pinne M, Rotondi ML, Miller MC, Demoll E, et al. Leptospira interrogans endostatin-like outer membrane proteins bind host fibronectin, laminin and regulators of complement. PLoS One (2007) 2(11):e1188. doi:10.1371/journal.pone.0001188
25. Choy HA, Kelley MM, Chen TL, Møller AK, Matsunaga J, Haake DA. Physiological osmotic induction of Leptospira interrogans adhesion: LigA and LigB bind extracellular matrix proteins and fibrinogen. Infect Immun (2007) 75(5):2441–50. doi:10.1128/IAI.01635-06
26. Barbosa AS, Monaris D, Silva LB, Morais ZM, Vasconcellos SA, Cianciarullo AM, et al. Functional characterization of LcpA, a surface-exposed protein of Leptospira spp. that binds the human complement regulator C4BP. Infect Immun (2010) 78(7):3207–16. doi:10.1128/IAI.00279-10
27. Murray GL. The molecular basis of leptospiral pathogenesis. In: Adler B, editor. Leptospira and Leptospirosis. Current Topics in Microbiology and Immunology. New York: Springer-Verlag Berlin Heidelberg (2015). p. 139–86.
28. Matsunaga J, Barocchi MA, Croda J, Young TA, Sanchez Y, Siqueira I, et al. Pathogenic Leptospira species express surface-exposed proteins belonging to the bacterial immunoglobulin superfamily. Mol Microbiol (2003) 49(4):929–45. doi:10.1046/j.1365-2958.2003.03619.x
29. Breda LC, Hsieh CL, Castiblanco Valencia MM, da Silva LB, Barbosa AS, Blom AM, et al. Fine mapping of the interaction between C4b-binding protein and outer membrane proteins LigA and LigB of pathogenic Leptospira interrogans. PLoS Negl Trop Dis (2015) 9(10):e0004192. doi:10.1371/journal.pntd.0004192
30. Choy HA. Multiple activities of LigB potentiate virulence of Leptospira interrogans: inhibition of alternative and classical pathways of complement. PLoS One (2012) 7(7):e41566. doi:10.1371/journal.pone.0041566
31. Castiblanco-Valencia MM, Fraga TR, Breda LC, Vasconcellos SA, Figueira CP, Picardeau M, et al. Acquisition of negative complement regulators by the saprophyte Leptospira biflexa expressing LigA or LigB confers enhanced survival in human serum. Immunol Lett (2016) 173:61–8. doi:10.1016/j.imlet.2016.03.005
32. Meri T, Amdahl H, Lehtinen MJ, Hyvärinen S, McDowell JV, Bhattacharjee A, et al. Microbes bind complement inhibitor factor H via a common site. PLoS Pathog (2013) 9(4):e1003308. doi:10.1371/journal.ppat.1003308
33. Souza NM, Vieira ML, Alves IJ, de Morais ZM, Vasconcellos SA, Nascimento AL. Lsa30, a novel adhesin of Leptospira interrogans binds human plasminogen and the complement regulator C4bp. Microb Pathog (2012) 53(3–4):125–34. doi:10.1016/j.micpath.2012.06.001
34. Siqueira GH, Atzingen MV, de Souza GO, Vasconcellos SA, Nascimento AL. Leptospira interrogans Lsa23 protein recruits plasminogen, factor H and C4BP from normal human serum and mediates C3b and C4b degradation. Microbiology (2016) 162(2):295–308. doi:10.1099/mic.0.000217
35. Wolff DG, Castiblanco-Valencia MM, Abe CM, Monaris D, Morais ZM, Souza GO, et al. Interaction of Leptospira elongation factor Tu with plasminogen and complement factor H: a metabolic leptospiral protein with moonlighting activities. PLoS One (2013) 8(11):e81818. doi:10.1371/journal.pone.0081818
36. Castiblanco-Valencia MM, Fraga TR, Pagotto AH, de Toledo Serrano SM, Abreu PA, Barbosa AS, et al. Plasmin cleaves fibrinogen and the human complement proteins C3b and C5 in the presence of Leptospira interrogans proteins: a new role of LigA and LigB in invasion and complement immune evasion. Immunobiology (2016) 221(5):679–89. doi:10.1016/j.imbio.2016.01.001
37. Raymond BB, Djordjevic S. Exploitation of plasmin(ogen) by bacterial pathogens of veterinary significance. Vet Microbiol (2015) 178(1–2):1–13. doi:10.1016/j.vetmic.2015.04.008
38. Sanderson-Smith ML, De Oliveira DM, Ranson M, McArthur JD. Bacterial plasminogen receptors: mediators of a multifaceted relationship. J Biomed Biotechnol (2012) 2012:272148. doi:10.1155/2012/272148
39. Bhattacharya S, Ploplis VA, Castellino FJ. Bacterial plasminogen receptors utilize host plasminogen system for effective invasion and dissemination. J Biomed Biotechnol (2012) 2012:482096. doi:10.1155/2012/482096
40. Vieira ML, Vasconcellos SA, Gonçales AP, de Morais ZM, Nascimento AL. Plasminogen acquisition and activation at the surface of Leptospira species lead to fibronectin degradation. Infect Immun (2009) 77(9):4092–101. doi:10.1128/IAI.00353-09
41. Verma A, Brissette CA, Bowman AA, Shah ST, Zipfel PF, Stevenson B. Leptospiral endostatin-like protein A is a bacterial cell surface receptor for human plasminogen. Infect Immun (2010) 78(5):2053–9. doi:10.1128/IAI.01282-09
42. Danø K, Andreasen PA, Grøndahl-Hansen J, Kristensen P, Nielsen LS, Skriver L. Plasminogen activators, tissue degradation, and cancer. Adv Cancer Res (1985) 44:139–266. doi:10.1016/S0065-230X(08)60028-7
43. Barthel D, Schindler S, Zipfel PF. Plasminogen is a complement inhibitor. J Biol Chem (2012) 287(22):18831–42. doi:10.1074/jbc.M111.323287
44. Vieira ML, de Morais ZM, Vasconcellos SA, Romero EC, Nascimento AL. In vitro evidence for immune evasion activity by human plasmin associated to pathogenic Leptospira interrogans. Microb Pathog (2011) 51(5):360–5. doi:10.1016/j.micpath.2011.06.008
45. Vieira ML, Atzingen MV, Oliveira TR, Oliveira R, Andrade DM, Vasconcellos SA, et al. In vitro identification of novel plasminogen-binding receptors of the pathogen Leptospira interrogans. PLoS One (2010) 5(6):e11259. doi:10.1371/journal.pone.0011259
46. Oliveira R, de Morais ZM, Gonçales AP, Romero EC, Vasconcellos SA, Nascimento AL. Characterization of novel OmpA-like protein of Leptospira interrogans that binds extracellular matrix molecules and plasminogen. PLoS One (2011) 6(7):e21962. doi:10.1371/journal.pone.0021962
47. Mendes RS, Von Atzingen M, de Morais ZM, Gonçales AP, Serrano SM, Asega AF, et al. The novel leptospiral surface adhesin Lsa20 binds laminin and human plasminogen and is probably expressed during infection. Infect Immun (2011) 79(11):4657–67. doi:10.1128/IAI.05583-11
48. Fernandes LG, Vieira ML, Kirchgatter K, Alves IJ, de Morais ZM, Vasconcellos SA, et al. OmpL1 is an extracellular matrix- and plasminogen-interacting protein of Leptospira spp. Infect Immun (2012) 80(10):3679–92. doi:10.1128/IAI.00474-12
49. Domingos RF, Vieira ML, Romero EC, Gonçales AP, Morais ZM, Vasconcellos SA, et al. Features of two proteins of Leptospira interrogans with potential role in host-pathogen interactions. BMC Microbiol (2012) 12:50. doi:10.1186/1471-2180-12-50
50. Nogueira SV, Backstedt BT, Smith AA, Qin JH, Wunder EA Jr, Ko A, et al. Leptospira interrogans enolase is secreted extracellularly and interacts with plasminogen. PLoS One (2013) 8(10):e78150. doi:10.1371/journal.pone.0078150
51. Fernandes LG, Vieira ML, Alves IJ, Morais ZM, Vasconcellos SA, Romero EC, et al. Functional and immunological evaluation of two novel proteins of Leptospira spp. Microbiology (2014) 160(Pt 1):149–64. doi:10.1099/mic.0.072074-0
52. Domingos RF, Fernandes LG, Romero EC, Morais ZM, Vasconcellos SA, Nascimento AL. Novel Leptospira interrogans protein Lsa32 is expressed during infection and binds laminin and plasminogen. Microbiology (2015) 161(Pt 4):851–64. doi:10.1099/mic.0.000041
53. Teixeira AF, de Morais ZM, Kirchgatter K, Romero EC, Vasconcellos SA, Nascimento AL. Features of two new proteins with OmpA-like domains identified in the genome sequences of Leptospira interrogans. PLoS One (2015) 10(4):e0122762. doi:10.1371/journal.pone.0122762
54. Silva LP, Fernandes LG, Vieira ML, de Souza GO, Heinemann MB, Vasconcellos SA, et al. Evaluation of two novel leptospiral proteins for their interaction with human host components. Pathog Dis (2016) 74(5). doi:10.1093/femspd/ftw040
55. Siqueira GH, Teixeira AF, Fernandes LG, de Souza GO, Kirchgatter K, Romero EC, et al. The recombinant LIC10508 is a plasma fibronectin, plasminogen, fibrinogen and C4BP-binding protein of Leptospira interrogans. Pathog Dis (2016) 74(2). doi:10.1093/femspd/ftv118
56. Cosate MR, Siqueira GH, de Souza GO, Vasconcellos SA, Nascimento AL. Mammalian cell entry (Mce) protein of Leptospira interrogans binds extracellular matrix components, plasminogen and β2 integrin. Microbiol Immunol (2016) 60(9):586–98. doi:10.1111/1348-0421.12406
57. Fraga TR, Courrol DS, Castiblanco-Valencia MM, Hirata IY, Vasconcellos SA, Juliano L, et al. Immune evasion by pathogenic Leptospira strains: the secretion of proteases that directly cleave complement proteins. J Infect Dis (2014) 209(6):876–86. doi:10.1093/infdis/jit569
58. Potempa M, Potempa J. Protease-dependent mechanisms of complement evasion by bacterial pathogens. Biol Chem (2012) 393(9):873–88. doi:10.1515/hsz-2012-0174
59. Potempa J, Pike RN. Corruption of innate immunity by bacterial proteases. J Innate Immun (2009) 1(2):70–87. doi:10.1159/000181144
Keywords: Leptospira, leptospirosis, complement system, immune evasion, serum resistance
Citation: Fraga TR, Isaac L and Barbosa AS (2016) Complement Evasion by Pathogenic Leptospira. Front. Immunol. 7:623. doi: 10.3389/fimmu.2016.00623
Received: 23 September 2016; Accepted: 08 December 2016;
Published: 21 December 2016
Edited by:
Melissa Jo Caimano, University of Connecticut Health Center, USAReviewed by:
Brian Stevenson, University of Kentucky, USAJames Matsunaga, University of California Los Angeles, USA
Elsio Augusto Wunder Jr., Yale University, USA
Copyright: © 2016 Fraga, Isaac and Barbosa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Angela Silva Barbosa, angela.barbosa@butantan.gov.br