





Abstract
Recent studies suggest a circannual pattern of bone turnover. To further investigate the underlying mechanisms, 41 healthy subjects (25‐80 years old) living in a southwestern German city were studied prospectively over a period of 18 months. Participants were examined every 4 weeks, and blood and urine samples were obtained on each visit. The following parameters were measured: serum 25‐hydroxyvitamin D3 [25(OH)D3], 1,25‐dihydroxyvitamin D3 [1,25(OH)2D3], and parathyroid hormone (PTH), as regulators, and serum total alkaline phosphatase (TAP), bone‐specific alkaline phosphatase (BAP), urinary total pyridinoline (PYD), deoxypyridinoline (DPD), and the aminoterminal telopeptide of collagen type I (NTX), as biochemical markers of bone turnover. The presence of significant circannual rhythms for the various markers was tested using the Pharmfit method. In the total group, 25(OH)D3, 1,25(OH)2D3, and PTH as well as BAP, PYD, DPD, and NTX showed a significant seasonal variation. 25(OH)D3 revealed the highest amplitude (38.0%) with an acrophase in August. Levels of the biochemical markers and of PTH were highest in winter with amplitudes of up to 17.7% (DPD). Results were most pronounced in premenopausal women, in subjects <50 years of age, and in subjects who did show a significant individual rhythm in 25(OH)D3 levels. No differences were found regarding other anthropometric or life style factors. Correlation analyses revealed strongest associations between the amplitudes of a vitamin D metabolite and a biochemical marker in premenopausal women. We conclude that specific markers of bone turnover show significant circannual rhythms. These changes are related directly to variations in the hormonal regulation of skeletal homeostasis. In postmenopausal women and in men, other effects may superimpose the circannual variation of biomarkers of bone turnover.
INTRODUCTION
ULTRADIAN, CIRCADIAN and infradian endocrine rhythms are considered crucial factors for the pathogenesis, prevention, and treatment of a wide range of diseases in almost all fields of medicine.(1) Skeletal homeostasis as an important metabolic system in the human organism also is subject to these naturally occurring oscillations. Thus, bone turnover underlies a significant circadian rhythm as indicated by the measurements of specific biochemical markers. (2–4) The menstrual cycle as an infradian rhythm also contributes significantly to variations in skeletal homeostasis.(5,6) In recent years, it has become evident that for bone turnover, biological rhythms of even longer periods, such as a circannual variation, may exist. (7–14) We and others have shown that bone turnover is accelerated during winter, but these reports are based mainly on cross‐sectional(7,8) or semilongitudinal(9,10) observations. In the few longitudinal studies, season‐related changes of bone turnover markers were less pronounced. However, the indices representing bone metabolism were rather unspecific (11–13) or the study population was relatively small.(14)
The importance of a circannual pattern of bone turnover is based, at least in part, on its potential relation to disturbances in skeletal homeostasis and the pathogenesis of chronic metabolic bone disease, that is, osteoporosis. Thus, it has been shown that both low bone mass and the rapidity of bone loss are independent predictors of future fracture risk. (15–17) A number of recent studies suggest that increased levels of bone turnover markers are associated with accelerated bone loss. (18–20) When exceeding the normal limits, a repeated seasonal acceleration of bone metabolism over the years therefore may contribute to the development of low bone mass and increase the risk for osteoporotic fractures.
Although there are few doubts that circannual fluctuations in bone turnover exist, the significance of this pattern is still controversial. To the best of our knowledge, a systematic evaluation of the known seasonal variation of osteotropic regulators (vitamin D and parathyroid hormone [PTH]) in the northern hemisphere (21–25) in relation to specific biochemical markers of bone turnover has not been reported yet. In the present longitudinal investigation of 41 volunteers followed over a period of 18 months we also evaluated the exact circannual pattern of biochemical parameters with different skeletal origin and the presence of cofactors potentially masking the effect of the season on bone turnover.
MATERIALS AND METHODS
Study population and data collection
A total number of 54 healthy volunteers aged 25‐80 years living in a suburban area of a southwestern German city (latitude, 49.5°N) initially were included in the present study. Of these, 15 men and 26 women completed the study and were followed over a period of 18 months starting in November 1996. None of the participants had evidence for the presence or a history of significant skeletal or nonskeletal disease or had taken any medication known to affect bone metabolism, including bisphosphonates, glucocorticoids, hormone‐replacement therapy (HRT), osteotropic vitamins, or calcium supplements. In addition, premenopausal women using oral contraceptives were excluded. Of the 26 women in the final analysis, 11 were postmenopausal with a mean duration of menopause (±SD) of 14.5 ± 10.5 years (median, 11.5 years; range, 1‐30 years). Thirteen of the 54 subjects had to be excluded from the final analysis because of incomplete data or voluntary study dropouts (n = 8), concurrent illnesses (n = 2), pregnancy (n = 1), and death (n = 2).
At study inclusion, a standardized interview was performed to obtain information on medical and reproductive history, as well as current and past use of medication. A questionnaire was applied to evaluate lifestyle habits, including alcohol intake, smoking, and physical activity. Weight and height measures were taken in light clothing without shoes. Obesity was estimated calculating the body mass index (BMI; kg/m2). Physical examination (i.e., blood pressure and heart rate) and evaluation of lifestyle factors were performed every 4 weeks. In premenopausal women, follow‐up visits took place on the same day of the respective menstrual cycle in order to minimize the influence of a potential variation of bone turnover markers caused by menstrual cycle‐related changes.
Nonfasting blood and urine samples were obtained during the same time of the day on each visit. To minimize laboratory imprecision and to assure that biochemical measurements were not affected by storage, the following procedure was applied. Blood samples were processed within 3 h after phlebotomy, centrifuged at 1500g for 10 minutes, and stored as aliquots at −80°C until analysis. Urine specimens were protected from light exposure and stored within 3 h of collection at −30°C until analysis. All laboratory analyses were performed within a total analytical period of 3 months after completion of patient recruitment and follow‐up. Assays were carried out in batches with all samples from a single patient run in one assay. The same batch of the respective assay was used for all measurements.
The study protocol was approved by the Ethics Committee of the University of Heidelberg, and written consent was obtained from all participants before inclusion.
Laboratory measurements
Serum 25‐hydroxyvitamin D3 [25(OH)D3, by radioimmunoassay; Incstar Corp., Stillwater, OK, U.S.A.], and serum PTH (by luminescence immunoassay; Ciba Corning Diagnostics, Walpole, MA, U.S.A.) were measured by commercially available immunoassays. The assay applied for the quantification of 25(OH)D3 detects the serum levels of both 25‐hydroxyvitamin D2 and 25(OH)D3. However, in contrast to the United States, there is no dietary vitamin D fortification in Germany. Therefore, the levels of 25‐hydroxyvitamin D2 detected by the assay are insignificant and contribute less than 1% to the total circulating vitamin D pool. 1,25(OH)2D3 was measured by in‐house radioimmunoassay as described previously.(26)
Bone formation markers:
Serum total alkaline phosphatase (TAP) was measured according to the optimized standard method of the “Deutsche Gesellschaft für Klinische Chemie,”(27) using an automated colorimetric assay (BM/Hitachi System 704 analyzer; Boehringer Mannheim GmbH, Mannheim, Germany) and p‐nitrophenyl phosphate as a substrate. Intra‐ and interassay coefficients of variation were <5%, with a normal range of 60‐170 U/liter in both sexes. A solid‐phase, two site immunoradiometric assay (Tandem‐Ostase; Hybritech Inc., San Diego, CA, U.S.A.) was applied to determine the serum concentration of bone‐specific alkaline phosphatase (BAP).(28) Intra‐ and interassay variations ranged between 3.7‐6.7% and 7.0‐8.1%, respectively. Normal values are 11.6 ± 4.1 μg/liter for premenopausal females and 12.4 ± 4.4 μg/liter for males.
Bone resorption markers:
Total urinary pyridinoline (PYD) and deoxypyridinoline (DPD) were determined by high‐performance liquid chromatography (HPLC). After complete acid hydrolysis of urine samples at 107°C for 16 h, the peptide‐free forms of PYD and DPD were separated by ion‐paired HPLC, and concentrations were quantitated by fluorometry using a fully automated method as described by Pratt et al.(29) Standards were derived from sheep bone and were a generous gift from Dr. Simon P. Robins (Aberdeen, Scotland). The overall reproducibility of the assay was between 3% and 12% for intra‐ and interassay variability. Urinary concentrations were expressed relatively to urinary creatinine (Cr) levels. Normal PYD values are 17.6‐47.3 nM/mM Cr (premenopausal females) and 10.8‐49.1 nM/mM Cr (males), and normal DPD values are 2.7‐12.7 nM/mM Cr (premenopausal females) and 2.6‐9.3 nM/mM Cr (males). Urinary concentrations of the immunoreactive aminoterminal telopeptide of collagen type I (NTX) were determined using a commercially available ELISA (Osteomark; Ostex International, Seattle, WA, U.S.A.) as described elsewhere.(30) Extinction was measured at 405 nm with a standard microplate reader performing all analyses in duplicate. Normal values are 51.0 ± 34.7 nM/mM Cr for premenopausal females and 69.4 ± 39.9 nM/mM Cr for males.
Other laboratory measurements:
All participants had routine laboratory evaluations, including the detection of serum and urinary Cr and total serum calcium (standardized colorimetric assays) using an automated method in a Beckmann II analyzer (Beckmann Instruments, Inc., Palo Alto, CA, U.S.A.). These measurements were performed immediately after phlebotomy and urine collections. Total serum calcium was corrected for total protein content.
Statistical analyses
The Statistical Analysis System (SAS) software package (SAS Institute, Cary, NC, U.S.A.) was used for descriptive and correlation analyses. Data analyses were performed for the total group and after stratification according to gender, age, menopausal status, BMI, 25(OH)D3 status, and lifestyle factors (smoking, alcohol intake, and physical activity). Descriptive statistics are presented as mean ± SD. The significance of group differences was tested using the Kruskal‐Wallis‐test or the Wilcoxon‐Mann‐Whitney U test, where appropriate. Fisher's exact test was used to test for differences in the distribution of participants within categories. Simple Pearson correlation coefficients and coefficients of determination were obtained to assess the strength of the association between the mesors or amplitudes of two markers.
Rhythm analyses were performed using the Pharmfit method as described previously.(31) The mesor (rhythm‐adjusted annual mean) and amplitude (half of peak‐to‐trough of rhythmic change) were calculated for each individual, assuming a period length of 365 days. Group data, expressed as percent of the individual mesor, were used for calculation of the acrophase (time of peak) and for rhythm statistics. The following equation was used for rhythm analyses: y = mesor + amplitude ∗︁ cos ([x − acrophase]/365 ∗︁ 2π), in which y is the measured marker value and x is the respective day of the year. Significance of circannual rhythmicity was tested using the zero‐amplitude hypothesis.
Univariate cofactor analyses were performed according to the following group stratifications: (1) males versus females, (2) males versus premenopausal females, (3) premenopausal versus postmenopausal females, (4) age <50 years versus age ≥50 years, (5) BMI <25.0 kg/m2 versus BMI ≥25.0 kg/m2, (6) physical activity <2 times per week versus physical activity ≥2 times per week, (7) alcohol consumption <2 times per week versus alcohol consumption ≥2 times per week, and (8) nonsmokers (including ex‐smokers and never smokers) versus smokers. Also, subjects who did show a significant individual seasonal rhythm in 25(OH)D3 [25(OH)D3+] were grouped together and analyzed as a separate group.
All statistical tests were two‐tailed and a significance level of p < 0.05 was considered statistically significant.
RESULTS
Circannual rhythms of bone turnover: total group
In the total group, serum 25(OH)D3, 1,25(OH)2D3, and PTH; serum BAP as a marker of bone formation; and urinary PYD, DPD, and NTX as bone resorption indices showed a significant seasonal rhythm as detected by Pharmfit analyses (Table 1). 25(OH)D3 revealed the highest amplitude with an acrophase in August. 1,25(OH)2D3 values resembled the pattern of 25(OH)D3, but the mean amplitude of the seasonal changes in 1,25(OH)2D3 was lower (17.5% of the mesor) compared with 38.0% of the mesor for 25(OH)D3.
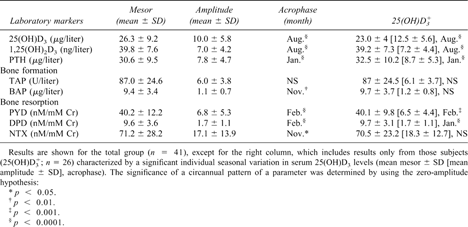
25(OH)D3 and 1,25(OH)2D3 showed an acrophase in August (group 1); BAP and NTX levels were highest in November (group 2); and peak values of PTH, PYD, and DPD were found in January/February (group 3). The acrophases of the markers within each of the three groups did not differ significantly. However, highly significant phase shifts were detected when comparing the acrophase of each marker in one group with the acrophase of each marker in the other two groups (p < 0.01 or greater, by Pharmfit analysis).
Circannual rhythms of bone turnover: cofactor analyses
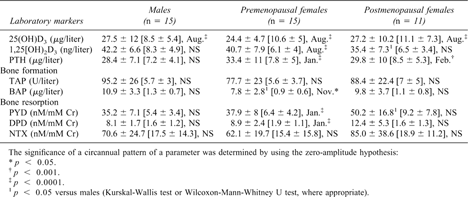
The purpose of the following analyses was to identify factors that may attenuate the seasonal pattern of markers of bone turnover and their association with vitamin D metabolites or PTH. Therefore, mean mesors and amplitudes of the applied parameters were analyzed first univariately according to the group stratifications as reported in the Materials and Methods section. Premenopausal women had on average significantly lower mesors of BAP than men (p < 0.05), and postmenopausal women showed a lower mean mesor of 1,25(OH)2D3 (p < 0.05) and a higher mean mesor of PYD (p < 0.05) than men. Otherwise, no significant group differences were detected (Table 2).
Circannual Rhythm of Hormonal and Biochemical Markers of Bone Turnover According to Gender and Menopausal Status (Mean Mesor ± SD [Mean Amplitude ± SD], Acrophase)
 |
|
Circannual Rhythm of Hormonal and Biochemical Markers of Bone Turnover According to Gender and Menopausal Status (Mean Mesor ± SD [Mean Amplitude ± SD], Acrophase)
|
|
In the next step, the seasonal patterns of markers of bone turnover were tested by Pharmfit analyses in each of the groups. Statistically significant differences in the calculated circannual rhythms for the various markers were found within the groups stratified according to gender, menopausal status (Table 2), and age (data not shown). The subclassification of the data set according to gender and menopausal status revealed most pronounced seasonal changes in vitamin D metabolites, PTH, and bone turnover markers in premenopausal females (Fig. 1). In this group and in subjects <50 years of age, a significant circannual variation was found for 25(OH)D3, 1,25(OH)2D3, and PTH, as well as for BAP, PYD, and DPD. In the male and postmenopausal female group as well as in older subjects (≥50 years), a significant rhythm was found for 25(OH)D3, 1,25(OH)2D3, and PTH only. None of the parameters studied showed a significant difference in acrophase between the groups. Other potential cofactors (i.e., BMI and lifestyle habits) were analyzed accordingly (see Materials and Methods section) but did not show any significant group differences.
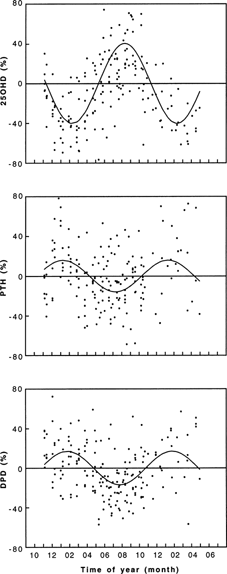
Circannual variation of hormonal and biochemical markers of bone turnover in premenopausal women, exemplified for 25(OH)D3, PTH, and DPD. Data are presented as individual values calculated as percentage of the respective mesor. The assessment of a circannual rhythm in marker values was performed using the Pharmfit method (see Materials and Methods section). The significance of a seasonal pattern was tested using the zero‐amplitude hypothesis.
Circannual rhythms of bone turnover: vitamin D status
None of the 41 participants who completed the study showed evidence of vitamin D deficiency. Thus, the individual mesors of 25(OH)D3 (range, 13.7‐60.9 μg/liter) and 1,25(OH)2D3 (range, 26.2‐57.1 ng/liter) were all within normal limits. On an individual basis, 63.4% of the participants showed a significant circannual rhythm in 25(OH)D3 levels. Because 25(OH)D3 served as a control parameter for season‐related changes, a further subclassification based on these results was performed. Mean mesors and amplitudes of vitamin D metabolites, PTH, and biochemical markers according to subjects who did show 25(OH)D3+, a significant individual rhythm in 25(OH)D3 levels, are presented in Table 1. In these subjects, a significant circannual variation was found for 25(OH)D3, 1,25(OH)2D3, and PTH, as well as for both urinary cross‐links.
Correlation analyses
Simple Pearson correlation analyses between the individual mesors or amplitudes of vitamin D metabolites, PTH, and biochemical markers of bone turnover were performed in the total and the stratified groups (see Materials and Methods section).
Overall, positive associations were found when correlating the mesors or the amplitudes of two respective biochemical markers of bone turnover in the total group or the various subgroups. Strongest and significant correlation coefficients were found between the three bone resorption markers in urine and between serum TAP and BAP (data not shown).
Overall, positive, but less pronounced correlations were found between mesors of the vitamin D metabolites and PTH. Highly significant and consistently positive associations were found for the amplitudes of 25(OH)D3 versus PTH throughout the total and the respective subgroups. The amplitude of 1,25(OH)2D3 was not correlated with the amplitude of 25(OH)D3 or PTH in any group (data not shown).
Special attention has been paid to potential associations between the individual mesors or amplitudes of a vitamin D metabolite or PTH and a biochemical marker of bone turnover. Of interest in this regard is the fact that in the subgroup with the most pronounced seasonal changes of bone turnover, that is, premenopausal women, no significant determination coefficients between the mesor of a vitamin D metabolite or PTH and a biochemical marker were found, except for a negative association of 25(OH)D3 versus TAP (r2 = 0.28; p < 0.05). However, in postmenopausal females, significant positive associations were found between the mesors of 25(OH)D3 and the urinary cross‐links [25(OH)D3 vs. PYD, r2 = 0.46 and p < 0.05; 25(OH)D3 vs. DPD, r2 = 0.40 and p < 0.05]. In contrast, overall positive associations between the amplitudes of 25(OH)D3 or 1,25(OH)2D3 and a biochemical marker were found in premenopausal [25(OH)D3 vs. BAP, r2 = 0.41 and p < 0.01; 25(OH)D3 vs. DPD, r2 = 0.32 and p < 0.05; 1,25(OH)2D3 vs. NTX, r2 = 0.31 and p < 0.05] and in postmenopausal women [1,25(OH)2D3 vs. NTX, r2 = 0.41 and p < 0.05; 1,25(OH)2D3 vs. DPD, r2 = 0.42 and p < 0.05]. No significant associations were found between the amplitudes of PTH and any of the biochemical markers in any of the groups.
DISCUSSION
The present study was carried out to systematically investigate previously suggested circannual changes of biochemical markers of bone turnover (7–14) and to evaluate their relation to changes in regulators of osteoblast and osteoclast function. To our knowledge, this is the first standardized longitudinal approach to address these questions using new and specific markers of bone turnover. Here, we show that all tested markers of collagen breakdown as well as the bone‐specific isozyme of AP indeed vary by season with increased levels in winter. These changes are accompanied by significant variations in serum 25(OH)D3, 1,25(OH)2D3, and PTH levels. However, no significant changes were found for serum TAP as a less specific marker of bone turnover.
The comparison of our data with previously published reports on seasonal variations of bone turnover(7–14, 32) requires consideration of the respective study settings (i.e., the latitude of the study location and other environmental factors), the study population (i.e., age and gender), the study design (i.e., longitudinal, cross‐sectional, or semilongitudinal), and the biochemical markers applied. Not surprisingly, in the present study, circannual changes of markers of bone turnover were most pronounced in premenopausal women. These subjects were selected and followed with regard to (i) a sufficient time interval from the pubertal growth spurt, (ii) the minimization of potential variations of bone turnover markers caused by the menstrual cycle, and (iii) the control of other potential cofactors. This strategy should generate a homogenous subpopulation in which the observation of subtle changes in bone turnover markers within the normal range is enabled. In this subgroup, we found significant circannual changes for BAP and both urinary cross‐links with acrophases in November and January, respectively. The seasonal variation of urinary NTX did not reach statistical significance. Because NTX is the marker with the highest assay variability,(33) larger numbers of subjects probably are required in order to establish a statistically significant circannual rhythm of NTX.
Four previous studies have investigated the seasonal effect on bone turnover markers in premenopausal women.(11, 13, 14, 32) However, in only one of these studies,(14) a clearly season‐related variation of a specific parameter of bone turnover, that is, serum osteocalcin, could be shown. The lack of seasonality in the other three studies(11, 13, 32) may be explained by either the young age (<25 years) of the participants(32) or the use of relatively unspecific biochemical markers, that is, TAP and hydroxyproline.(11,13) Considering the latter, our data clearly confirm the lack of specificity of serum TAP,(28) as a significant seasonal variation could not be detected for this marker.
The circannual pattern of bone turnover marker levels was similar in pre‐ and postmenopausal women and in men, although statistically significant changes could not be observed in the latter two groups. Notably, we and others have previously reported significant seasonal variations in markers of bone turnover in postmenopausal women and in men. (7–10) In contrast to the present investigation, these studies have been performed as cross‐sectional or semilongitudinal evaluations in large cohorts. Compared with these previous reports and given the similar pattern of the circannual variation of bone turnover markers in the three subgroups, a seasonality of bone turnover in men and in postmenopausal women also is strongly suggested. However, powerful covariates such as the overall increased bone turnover caused by estrogen deficiency in postmenopausal women(34) or increased morbidity(35) may superimpose the physiological season‐related changes in marker levels in these subgroups. It also should be noted that, although the study design was longitudinal and power analyses revealed a sufficient number of data points for Pharmfit‐based rhythm analyses even in the smaller subgroups, the sample sizes are still small. Therefore, false‐negative results are not ruled out completely for biomarkers with relatively small circannual changes in these subgroups, and the lack of seasonality of some markers may have to do with statistical power.
It is believed that anthropometric variables, for example, age(36) or body mass,(37) and lifestyle factors such as physical activity,(38) smoking,(39) or alcohol intake,(40) exert an effect on bone turnover that may be reflected by the measurement of specific biochemical markers. These factors as well as the assay variability of marker measurements(33) may well attenuate the circannual variation of bone turnover markers.(7–14, 32) We therefore performed subanalyses according to the group classifications outlined in the Material and Methods section. However, no significant differences in the circannual pattern of bone marker levels were found in the various groups, except for a subclassification according to age. As in postmenopausal women and in men, the less pronounced seasonality of bone turnover in participants >50 years of age is most probably caused by powerful covariates.
It has often been difficult to associate directly the well‐known seasonal changes in 25(OH)D3 levels with the fluctuations in levels of biochemical markers of bone turnover.(7, 11, 32) Accordingly, in the present study we did not find consistent correlations of the mesors of a vitamin D metabolite or PTH with a biochemical marker. (The mesor as the annual mean of a respective marker in an individual participant is comparable with absolute marker levels used for correlation analyses in cross‐sectional investigations.) We suggest that at least part of these results may be caused by the study design. Thus, in the present healthy population, the fluctuations of osteotropic regulators within the normal range may not exert a strong effect on biochemical marker levels. However, despite the poor correlation coefficients between the mesors, we indeed showed a direct relationship between the seasonal changes of the vitamin D status and bone turnover.
First, the fluctuations of the biologically inert 25(OH)D3 as the major circulating form of vitamin D were paralleled by a less pronounced, but still highly significant seasonal variation in 1,25(OH)2D3 levels. The latter is the most important vitamin D metabolite for actions on bone and calcium homeostasis.(41) It is easily conceivable that changes in serum concentrations of 1,25(OH)2D3 cause direct effects on bone cells and, thus, bone turnover.
Also, most pronounced in premenopausal women, we found significant correlation coefficients between the amplitudes of 25(OH)D3 with BAP and DPD as highly specific markers of bone turnover. Clearly, in the present healthy study population, the distribution of the amplitudes most likely reflects the normal profile of circannual changes of biochemical markers of bone turnover. However, we suggest the presence of a threshold for vitamin D effects on changes in marker values. Thus, a person with an otherwise normal mesor of 25(OH)D3, and although not frankly 25(OH)D3‐deficient, may fall under the 25(OH)D3 threshold at certain time points during the study period. Lower‐than‐threshold 25(OH)D3 values may indicate a state of latent vitamin D deficiency at which the skeletal homeostasis is already deranged. This situation may lead to increased levels of specific markers of bone turnover and could be reflected by the highly positive correlations of the amplitudes of 25(OH)D3 and the bone markers. Our hypothesis is supported by the fact that in patients with secondary hyperparathyroidism and, thus, the presence of long‐term lower‐than‐threshold 25(OH)D3 levels, 25(OH)D3 is indeed significantly correlated with bone marker levels.(8)
The biological significance of seasonal changes in bone turnover has yet to be determined. However, it is obvious that the circannual variations of biochemical markers have to be considered when interpreting laboratory results. If bone marker levels reflect the actual rate of bone turnover, our findings also may be relevant in the pathophysiology of metabolic bone disease. It has been shown clearly that high levels of bone markers are associated with rapid bone loss. (18–20) Moreover, increased marker values are independent risk factors for the development of osteoporotic fractures.(42,43) Thus, accelerated bone turnover in winter, if exceeding normal limits, over the years may contribute to the reduction of bone mass. A role for seasonal influences on bone metabolism also is suggested by the fact that the incidence of osteoporotic fractures is higher in populations of the northern hemisphere, regardless of ethnic differences.(44) Given the results for premenopausal women, our study may indicate that an early onset of vitamin D supplementation in winter is of benefit in terms of the long‐term prevention of bone loss. However, preceding such a definitive recommendation, the season‐related association between bone mass and bone turnover has to be clearly demonstrated, although data on seasonal changes of bone density measurements with lower values in winter have been published.(9, 12, 45) Moreover, controlled intervention studies, including the supplementation of vitamin D in winter, have to be performed.
ACKNOWLEDGMENTS
We thank Beatrice Auler, Christian Kissling, Antje von Schickfuss, and Uschi Woitge for excellent technical assistance. We are grateful to Dr. Gerhard Wetzig, Lindenfels, Germany, for providing the local equipment to process blood samples immediately. This work was supported by a grant from the University of Heidelberg (F203382) to H.W.W.

{kind=link}
{kind=link}
{kind=link}