
Abstract
The musculoskeletal system is a tight network of many tissues. Coordinated interplay at a biochemical level between tissues is essential for development and repair. Traumatic injury usually affects several tissues and represents a large challenge in clinical settings. The current demand for potent growth factors in such applications thus accompanies the keen interest in molecular mechanisms and orchestration of tissue formation. Of special interest are multitasking growth factors that act as signals in a variety of cell types, both in a paracrine and in an autocrine manner, thereby inducing cell differentiation and coordinating not only tissue assembly at specific sites but also maturation and homeostasis. We concentrate here on bone morphogenetic proteins (BMPs), which are important crosstalk mediators known for their irreplaceable roles in vertebrate development. The molecular crosstalk during embryonic musculoskeletal tissue formation is recapitulated in adult repair. BMPs act at different levels from the initiation to maturation of newly formed tissue. Interestingly, this is influenced by the spatiotemporal expression of different BMPs, their receptors and co-factors at the site of repair. Thus, the regenerative potential of BMPs needs to be evaluated in the context of highly connected tissues such as muscle and bone and might indeed be different in more poorly connected tissues such as cartilage. This highlights the need for an understanding of BMP signaling across tissues in order to eventually improve BMP regenerative potential in clinical applications. In this review, the distinct members of the BMP family and their individual contribution to musculoskeletal tissue repair are summarized by focusing on their paracrine and autocrine functions.
Similar content being viewed by others

Introducing the BMP pathways: from single function to musculoskeletal tissue crosstalk
Bone morphogenetic proteins (BMPs) and growth and differentiation factors (GDFs) are members of the transforming growth factor beta (TGFβ) superfamily of cytokines. Today about 20 different BMPs with redundant, non-redundant, or even opposing cellular functions, including the regulation of cell differentiation, proliferation, survival, migration and homeostasis, are known. BMPs are secreted growth factors and are commonly categorized on the basis of their structural properties; they have been extensively studied in the past with a focus on their differentiation potential with regard to bone precursor cells (Miyazono et al. 2005). However, ongoing research has revealed that BMPs exert pleiotropic effects in tissues other than bone and are clearly multitasking mediators rather than classical morphogens with a single function (Wagner et al. 2010; Reddi 2005). In this review, we focus on their roles in the musculoskeletal system, which is highly connected but, at the same time, structurally distinct. During repair processes of all musculoskeletal tissues, the mediating and multitasking role of BMPs becomes strikingly visible. Here, we briefly introduce the BMP pathway components and associated signaling cascades that are described in detail in the individual sections of this review.
Amongst all known BMPs, we discuss BMP2-4, BMP6, BMP7, BMP9 and BMP12-14. They differ in their expression, their affinities for BMP receptors, and their extracellular antagonists and fine-tuners such as Noggin, Gremlin and BMP-binding endothelial-cell-precursor-derived regulator (BMPER; Krause et al. 2011; Stabile et al. 2007; Heinke et al. 2008). They also differ in their availability for a certain tissue because of their specific binding to extracellular matrix (ECM) components (Seib et al. 2010; Shah et al. 1999). The activity and specific function of certain BMPs thus relies not only on the coordinated inhibition by antagonists but also on the tissue availability and local presence of the cognate cell surface receptors. Active BMPs bind to transmembrane serine/threonine kinase receptors that form heteromeric complexes composed of type I and type II receptors (BMPRI and BMPRII). Type I receptors include activin receptor type Ia (ACTRIa), BMPRIa and BMPRIb. Type II receptors comprise BMPRII and activin receptor type IIa (ACTRIIa) and ACTRIIb (Nishitoh et al. 1996). Additional signal regulation is achieved by tissue specific co-receptors such as Endoglin, Ror2 and repulsive guidance molecules (RGMs; Barbara et al. 1999; Sammar et al. 2004; Halbrooks et al. 2007). The mechanisms by which BMPs activate intracellular signaling are best characterized for BMP2. BMP2 binding to a preformed complex (PFC) of type I and type II receptors initiates Smad signaling (Fig. 1a). Additionally, when BMP2 binds to its high affinity type I receptor, the type II receptor is recruited into the complex forming the so-called BMP-induced signaling complex (BISC). After ligand binding and receptor-complex formation, the constitutive active type II receptor transphosphorylates the type I receptor at the GS box located at the juxtamembrane part of the receptor. The activated type I receptor in turn phosphorylates receptor-bound Smads1/5/8 (R-Smads) at the C-terminal Mad homology 2 (MH2) domain leading to conformational changes and subsequent release into the cytoplasm. Activated Smads assemble with Common mediator Smad 4 (Co-Smad) and are translocated into the nucleus where they induce specific target gene transcription via binding of co-factors and Smad-responsive DNA elements (Chen and Massague 1999; Massague et al. 2005; Sieber et al. 2009).

Representation of signaling pathways activated by bone morphogenetic protein (BMP) receptor (BMPR) complex. Upon BMP binding to the heterotetrameric serine/threonine kinase complex of BMP receptor type 1 (BMPRI) and BMPRII, the phosphorylation of BMPRI-bound Smads1/5/8 (R-Smads) is initiated leading to a quick release into the cytoplasm. There, phosphorylated Smads oligomerize with co-Smad 4 and further co-factors and the whole complex is finally translocated into the nucleus where it induces Smad-dependent target genes (SBE SMAD-binding element; a). BMPs also induce pathways not directly linked to the functions of Smads; these are referred to as non-Smad signaling. Phosphoinositide-3-kinase (PI3K) is activated by BMPs and regulates the activity of the survival kinase Akt and small Rho GTPases, which are key mediators in filamentous actin (F-actin) formation. LIM domain kinase (LIMK) associates with BMPRII and regulates the activity of cofilin, an actin-binding protein. The X-linked inhibitor of apoptosis protein (XIAP) binds to BMP receptors and recruits transforming-growth-factor-β-activated kinase1 (TAK) and TAK-binding protein (TAB) into a complex to initiate the BMP-induced mitogen-activated protein (MAP) kinase (MAPK) pathway. Activated MAPK induces both migration and transcriptional responses. Furthermore, MAPK crosstalks to the Smad pathway by phosphorylation of the linker region of Smads to prime Smads for ubiquitination and degradation (b)
The BISCs activate the p38 mitogen-activated protein kinase (p38 MAPK) and thereby initiate non-Smad signaling responses (Fig. 1b; Mu et al. 2011). This mode of signaling is characterized by the recruitment of the X-linked inhibitor of apoptosis (XIAP), TGFβ-activated kinase 1 (TAK1) and TAK-binding protein (TAB; Yamaguchi et al. 1999; Sorrentino et al. 2008; Liu et al. 2009). This adaptor complex regulates the mitogen-activated kinase kinases (MKK) leading to activated MAPKs. In addition to p38 MAPK, a BMP-dependent activation of extracellular regulated kinase 1,2 (Erk1,2) and c-Jun-NH2-terminal kinase (JNK)/stress-activated protein kinase (SAPK) have been reported (Zanotti et al. 2008; Guicheux et al. 2003). Beside the Smad and MAPK cascades other non-Smad signaling pathways are in the focus of current investigations since they contribute to the pleiotropic effects of BMPs. Phosphoinositide-3-kinase (PI3K) is activated by BMPs and generates the membrane-bound second messenger phosphatidylinositol 3,4,5-tri-phosphate (PIP3; Ghosh-Choudhury et al. 2002). PIP3 regulate the activity of proteins involved in cell survival and migration, such as the survival kinase Akt/PKB or the family of small Rho family GTPases (Rho GTPases), respectively (Cain and Ridley 2009).
Whereas Akt signaling has been shown to be important for BMP-dependent differentiation, RhoGTPases are crucial players in the rearrangement of the actin cytoskeleton mediating the formation of filamentous actin (F-actin; Hall 1998). Another BMP-regulated kinase and key player of cytoskeletal rearrangements is the Lim domain kinase (LIMK; Foletta et al. 2003). LIMK has been shown to rearrange the actin cytoskeleton via actin depolymerizing factor (ADF)/cofilin pathways in a BMP-dependent fashion (Soosairajah et al. 2005). P38 MAPK activation has recently been demonstrated to influence BMP-mediated actin remodeling via the activation of heat shock protein 25 (HSP25) in addition to its role in transcriptional regulation of ALP (Gamell et al. 2011).
In summary, BMP signaling is tightly controlled and induces several cellular responses including transcriptional and non-transcriptional events. These become additionally influenced by a multitude of regulatory factors defining signaling intensity, duration and efficacy. This pattern of complexity is recapitulated in musculoskeletal tissue regeneration involving autocrine processes that induce one or more cellular responses in a single tissue and paracrine processes that occur between two or even more tissues and that we term “tissue crosstalk”. Because a high regeneration potential is linked to the tight interaction of tissues, we review here the known BMP crosstalk mechanisms in a hierarchical manner starting from the highly connected tissue skeletal muscle-tendon-bone to the less connected cartilage. We concomitantly indicate the importance of the co-regenerating vascular and nervous systems.
BMP signaling in muscle regeneration
The skeletal muscle represents the perfect tissue to start this overview of tissue crosstalk during regenerative processes because it is highly connected to bones by bundles of collagen fibers known as tendons. This connection exists not only in a mechanical form but also through several signaling cascades. Regenerative signaling pathways in the skeletal muscle are thought to recapitulate the main processes that also occur during embryonic muscle development. However, in some aspects, muscle regeneration seems to act differently, since the migration of adult myogenic precursor cells (satellite cells) is not needed as compared with the embryonal migration of somite-derived cells to their target site (Birchmeier and Brohmann 2000; Brohmann et al. 2000). Upon muscle injury, growth factors secreted by cells of the immune system accumulate within the ECM around lesions; this is the priming event for satellite cell activation and subsequent muscle regeneration (Carlson 2003; Husmann et al. 1996). The paired box protein 7 (Pax7) is the best characterized satellite cell marker. Pax7 −/− mice display normal muscle development but lack satellite cells in adult muscle tissue (Seale et al. 2000). This phenotype implies the different origin of embryonic muscle precursors versus satellite cells. Pax7 deficiency during embryonic muscle development is compensated by other members of the Pax family (e.g., Pax3), whereas Pax7 is essential for satellite cell viability during adult stages. After injury, activated Pax7-positive cells turn into a proliferative form giving rise to fusion-competent myoblasts expressing the early muscle-specific determination factor Myf5 (Kuang et al. 2008; Rudnicki et al. 2008). These myoblasts are able to fuse with damaged muscle fibers or with themselves to generate new functional muscle tissue (Fig. 2b). A small population of myoblasts does not express Myf5 and retains the quiescent satellite cell character to repopulate the stem cell niche within the muscle (Kuang et al. 2007).

Role of BMPs in tendon-bone, tendon-muscle and tendon-tendon interplay. Scleraxis (SCX)-driven BMP4 secretion from growing tendons induces bone ridge formation by activating chondrocytes via BMPRIa and BMPRII (a). Tendons also crosstalk to injured muscle tissue via paracrine BMP4 secretion acting on muscle-located paired box protein 7 (Pax7)-positive satellite progenitor cells. Those respond by proliferation via BMPRIa, BMPRII and inhibitor of differentiation (Id) proteins 1 and 3. Following myoblast formation, repair processes become terminated by increasing amounts of noggin. In addition to proliferating satellite cells, a BMP-nonresponsive and quiescent satellite cell population can be found (b). Tendon regeneration is also mediated by BMPs. Here, mesenchymal progenitor cells have been shown to differentiate in a SXC- and Smad8-dependent manner, whereas BMP12-14 have been demonstrated to be important regulators of tenocyte proliferation and differentiation (c)
Several regulatory pathways control satellite cell functions, the most prominent being Delta/Notch and Wnt signaling (Wagers and Conboy 2005; Zammit 2008). BMPs are known to inhibit myogenic differentiation during embryogenesis in order to promote osteogenic differentiation (Amthor et al. 1998; Reshef et al. 1998). However, a BMP-dependent pro-proliferative function on satellite cells is found during adult muscle regeneration, when BMP signaling drives satellite cell proliferation and prevents their differentiation through the inhibition of muscle differentiation factor MyoD. Proliferating satellite cells from murine muscle fibers have been shown to express high levels of BMPRIa (ALK3), activated Smad1/5/8 and Smad4. Administration of BMP4 to mice with induced muscle damage sustains satellite cell division and reduces differentiation, whereas the blocking of BMP signaling induces premature differentiation. Intervention with BMP signaling has been achieved by using the BMP antagonist Noggin, BMPRIa knockdown, or soluble BMPRIa receptor (ligand trap; Ono et al. 2011).
BMP4 is secreted by myogenic precursor cells. For example, the pluripotent muscle progenitor cell line C2C12 expresses and secretes the ligand in marginal concentrations under low serum conditions (Clever et al. 2010). The secreted BMP4 is functional and induces the expression of BMP targets, namely inhibitor of differentiation 1 (Id1) and Id3, whereas Noggin significantly decreases pSmad1/5/8 and the expression levels of Id1 and Id3. With regard to the in vitro osteoinductive potential of BMP4 on C2C12 cells, cell fate decisions seem to be highly dependent on ligand concentrations. BMP4 expression has also been observed in quiescent satellite cells (CSM4B cells) isolated from 2-month-old mice (Kumar et al. 2009) and in human skeletal muscle. Increased myoblastic BMP4 expression might be responsible for the progressive muscle weakness and ineffective muscle regeneration in patients suffering from Duchenne muscular dystrophy (DMD). Primary myoblasts from skeletal muscle biopsies of DMD patients express about three-fold more BMP4 compared to healthy persons; this increase is suggested to be responsible for the less effective differentiation of myoblasts into myotubes in DMD patients (Sterrenburg et al. 2006). In addition to myogenic progenitors, another BMP source is represented by immune cells, such as macrophages, which are recruited to the damaged muscle as a result of inflammation (Xiao et al. 2010). Circulating BMP4 might be also delivered directly to the injured site through ruptured vessels. These muscular hemorrhages contribute to stimulate satellite cells within the injured muscle through BMP4 circulating in a large 100-kDa complex in the blood stream (Kodaira et al. 2006). Interestingly, satellite cells are localized close to blood vessels and their number positively correlates with the capillarization of the myofiber, indicating the occurrence of a blood vessel/satellite cell crosstalk that regulates myogenic cell expansion, myogenesis and angiogenesis (Mounier et al. 2011).
Crosstalk between muscle and tendon
Transmission of the contractile force produced by skeletal muscle fibers from myofilaments to tendon fibers occurs at the muscle-tendon junction. BMP-dependent mechanisms regulate the development of this tissue interaction site during fetal myogenesis (Edom-Vovard and Duprez 2004). In developing chick limbs, a subpopulation of Pax7-positive and BMP-responsive cells has been identified at the tips of the muscle in close proximity to tendon tissue producing BMP4. This paracrine BMP signal regulates the number of progenitors and satellite cells during myogenesis to maintain their proliferation (Fig. 2b). Actively dividing Pax7-positive cells display Smad1/5/8 activation and have been found in clusters located at the tips of isolated adult muscle fibers. Ectopic Noggin expression reduces the number of these progenitors and fibers resulting in smaller muscles (H. Wang et al. 2010). The specific location of BMP-responsive satellite cells agrees with the observations of myogenic progenitor accumulation at the ends of growing skeletal muscle fibers of the post-hatch chicken (Allouh et al. 2008) and increased satellite cell proliferation close to the tendons (Tsujimura et al. 2006). In adult myofibers, two populations of satellite cells are found: a quiescent BMP-signaling-independent subgroup and a BMP-responsive proliferating subgroup contributing to muscle repair. These findings in combination with the observation that BMP4 originates from tenocytes suggest a leading role of BMPs in muscle-tendon crosstalk during regeneration processes at adult stages. Thus, tendons clearly play a central role not only as a physical linker between muscles and bone but also as a signal-mediating tissue.
BMP signaling in tendon
Tendons and ligaments are fibro-elastic structures mainly composed of collagen types I and III, elastin, proteoglycans and structural glycoproteins, connecting muscles with bones to communicate muscle motor activity to the bone for proper joint movement. Tendon fibroblasts, called tenoblasts and/or differentiated tenocytes, represent about 90% of the cellular components of the tendon. The cells are aligned into longitudinal rows with numerous sheet-like structures. Cell-cell communication is provided by gap junctions between tenocytes within the same row and between cells of adjacent rows (Waggett et al. 2006). In addition to structural components, only a few molecules have been described as general tendon markers (Edom-Vovard and Duprez 2004): scleraxis (SCX; Schweitzer et al. 2001), tenascin (Ros et al. 1995), fibroblast growth factor 8 (Edom-Vovard et al. 2001) and BMP4 (D’Souza and Patel 1999). The cell-to-matrix ratio decreases during life resulting in morphological and metabolic changes of the tendon cells (Kannus 2000). In addition to the more than 30 million tendon and ligament injuries reported globally each year (Maffulli et al. 2003), changes in the metabolic activity can lead to pathological patterns in tendon structure caused by the overproduction and/or oversecretion of degradative enzymes or by matrix loss through cellular phagocytosis and pinocytosis comparable with the activity of osteoclasts in bone resorption (Jozsa et al. 1990). In vivo studies on tendon regeneration have to be carefully distinguished between tendon-tendon studies and experiments performed on the muscle-tendon or tendon-bone interfaces. In the following, the current literature according to tendon-tendon studies and studies on the tendon-bone junction will be outlined in more detail. Crosstalk between chondrocytes/osteocytes and tenocytes has an important impact on the quality of newly formed tendon, especially in the context of BMP action. For example, for the effective healing of the tendon-bone interface, stimuli that address both tenocytes with their slow metabolism and the more active osteoblasts obviously have to be used. Several growth factors such as insulin-like-growth-factor-1 (IGF1), platelet-derived growth factor, TGFβ, or BMPs have been described to exert positive effects on the proliferation and differentiation of tenocyte progenitors (X.T. Wang et al. 2004; Violini et al. 2009; Tsubone et al. 2004; Dahlgren et al. 2005; Wong et al. 2005; Pauly et al. 2011; Lou et al. 2001).
BMPs in neotendon formation and repair
The way that mesenchymal progenitors are efficiently programmed to yield several cell lineages, e.g., chondrocytes, osteocytes, myocytes and tenocytes, is one of the major research fields for musculoskeletal tissue engineering. C3H10T1/2 cells are a murine multipotent mesenchymal cell line showing a similar differentiation potential to paraxial mesenchymal progenitors in response to distinct stimuli. The ectopic expression of recombinant BMP2, for example, induces osteogenesis, whereas the application of BMP12 or BMP13 leads to tenogenesis (Fig. 2c). All three ligands show similar affinity to the BMP type I receptors BMPRIa and BMPRIb and to the type II receptors ACTRIIa, ACTRIIb and BMPRII. Treatment of C3H10T1/2 with recombinant BMP2 induces Smad phosphorylation and the expression of osteogenic markers such as osteocalcin, whereas recombinant human (rh) BMP12 or BMP13 administration results in the expression of specific tenogenic genes such as thrombospondin 4 without the activation of Smads (Berasi et al. 2011); this clearly demonstrates that the activation of the same receptors by different BMPs activates both Smad and non-Smad signaling to enable manifold differentiation processes. The current literature emphasizes Smad8 as a tendon-specific Smad; however, the way that the mechanism of action is regulated remains unclear. A special setting of BMP2 overexpression and co-expression of biologically active Smad8 (∆MH1) in C3H10T1/2 cells has been shown to influence neotendon formation (Hoffmann et al. 2006; Towler and Gelberman 2006). Tendon-specific cell differentiation in vitro with the induction of tendon-specific genes such as Scx, Six1 and 2, EphrinA4 and ProcollagenI has been observed and confirmed in vivo through the implantation of these cells seeded on collagen sponges into an Achilles-tendon-defect model in rats. Neither Smad1 nor Smad5, which lack the MH1 domain, are able to induce tenogenic differentiation, suggesting a specific role of Smad8 in tendon formation. Conflicting data have been published regarding the cartilage- or tendon-inducing effect of GDF implants. GDF5 (BMP14, CDMP1), GDF6 (BMP13, CDMP2) and GDF7 (BMP12, CDMP3) have the potential to induce the ectopic formation of collagen type I fibers when injected subcutaneously or intramuscularly (Wolfman et al. 1997). Histological analysis has disclosed a wave-form of fibers with regular periodicity resembling tendon and ligament morphology. Malformation in tendon pattering has been described in GDF5 knockout mice (bp/bp mice), which show skeletal and connective tissue deformations at sites of the tendon-bone interfaces, indicating linked mechanisms between the formation of bone and tendon structures depending on specific time- and dosage-dependent growth factor induction (Clark et al. 2001; Gruneberg and Lee 1973; Mikic 2004). After Achilles tenotomy, GDF5-deficient mice show a 1- to 2-week delay in tendon healing with decreased cell density and collagen content and a 1-week delay in angiogenesis of the newly built tissue (Chhabra et al. 2003). GDF6 (BMP13) knockout mice, however, show gender-dependent structural and functional tendon-specific malformations characterized by reduced collagen and glucosaminoglycan content in tail-tendon fascicles with compromised material properties in the male only (Mikic et al. 2009a, 2009b). BMP12 seems to be one of the most potent tenogenic growth factors of the BMP family as shown in several in vitro differentiation studies of mesenchymal stem cells (MSCs). BMP12 administration to adult MSCs reproducibly and significantly induces the expression of tendon marker genes such as Collagen type I, SCX and Tenomodulin (Q.W. Wang et al. 2005; Lee et al. 2011). Application of recombinant BMP12 increases the proliferation of tendon fibroblasts in a dose-dependent manner and the expression of typical extracellular tendon compounds such as procollagen types I and III. These findings indicate the importance of BMP12 in matrix remodeling processes and early phases of tendon regeneration in adult tissues (Fu et al. 2003).
Crosstalk at the tendon-bone interface
BMPs regulate both longitudinal bone growth and patterning processes leading to the bone ridge structure. The latter is important for tendon-skeleton interactions (Fig. 2a). Bone ridges ensure the functionality of the musculoskeletal system, providing an anchoring point for skeletal muscle via tendon attachment. BMP4 has been shown to be the most important BMP mediating crosstalk between tendon and bone, being necessary for the initiation of bone ridge development in a biphasic process (Blitz et al. 2009). SCX, a tendon marker expressed in progenitors and cells of all tendonous tissues, regulates tenoblast differentiation and acts upstream of BMP4 expression in tendon cells. In Scx −/− embryos, BMP4 expression is lost at the tendon insertion site. In the second step, secreted BMP4 originating from the tendons binds to adjacent chondrocytes via BMPRIa/BMPRII complexes, which in turn activates a BMP signaling cascade that subsequently induces bone ridge formation.
Furthermore, BMP2, BMP4 and BMP7 have an osteoinductive influence as shown in the ectopic chondro-ossification of regenerating tendons resulting in failed tendon healing (Yee Lui et al. 2011). Retroviral delivery of Noggin or the application of muscle-derived stem cells overexpressing Noggin are able to inhibit the BMP4-induced heterotropic ossification of tendon (Glaser et al. 2003; Hannallah et al. 2004). The precise spatiotemporal application of specific BMP pathway inhibitors might be an interesting strategy to improve tendon healing and to counterbalance unwanted ossification processes.
Within the BMP family, BMP2 and BMP7 are well known for their ability to induce bone and cartilage formation by regulating the differentiation of mesenchymal progenitor cells (E.A. Wang et al. 1990; Fang et al. 1996), whereas only a few studies have commented on the positive effects of BMP2 and BMP7 with regard to the regeneration of the tendon-bone interface. Both tenocytes and osteoblasts, isolated from human rotator cuff tissue, express high affinity receptors for BMP2 and BMP7, whereas tenocytes show lower receptor expression levels (Pauly et al. 2011). By using a tendon-bone healing ovine model, BMP7 expression has been detected at the tendon-bone interface close to the bony side, whereas BMP2 expression shifts to the tendon side at 6 weeks after injury (Yu et al. 2007). Other data imply a positive effect of BMP2 application on previously regenerated tendon-bone borders by improving the collagen type I production in tenocytes and/or osteoblasts (Martinek et al. 2002; Kohno et al. 2007). BMP7 triggers the production of fibrous tissue between tendon and bone comparable with native regeneration processes in the tissue-connecting area (Rodeo et al. 1993; Gerber et al. 1999). BMP7 also increases the proliferation and expression of tenocyte markers such as CollagenI and proteoglycans in a dose-dependent manner (Yamada et al. 2008). In gene therapy studies of BMP2-overexpressing rat cells embedded in calcium alginate gels, a positive effect on tendon-bone healing has been observed in rabbits after anterior cruciate ligament reconstructions. Concomitantly, increased expression of the vascular endothelial growth factor (VEGF) promotes angiogenesis (C.J. Wang et al. 2010), which together with the improved tendon-bone integration is desired in late tendon and bone healing processes. These crosstalk mechanisms between the bone and vascular system will be described under Crosstalk between endothelium and bone.
BMPs in bone regeneration
Here, we will outline the bone healing process that is highly regulated by BMPs. In contrast to other tissues, bone is characterized by a high regeneration potential controlled by bone itself and by interplay with factors from the immune system and the vascular system. The latter system is rebuilt in parallel during bone healing and will be described in detail under Crosstalk between endothelium and bone. However, 5%–10% of fractures are associated with delayed healing or non-unions, often caused by a limited blood supply. Today, the treatment of non-unions is achieved by non-surgical ultrasound or pulsed electromagnetic stimulation and by surgical intervention by using bone grafts, bone graft substitutes, or internal/external fixation, all with the optional addition of rhBMP2 or rhBMP7 (White et al. 2007; Einhorn et al. 2003; Carlisle and Fischgrund 2005). The outstanding role of BMPs in bone regeneration has further been investigated in preclinical studies in which the in vitro and the in vivo gene transfer of BMPs have been shown to enhance healing (Nauth et al. 2010).
Postnatal bone regeneration is a complex series of overlapping processes that recapitulate embryonic skeletal development. During fracture healing, bone is formed by endochondral and intramembranous ossification. Endochondral bone formation occurs external to the periosteum in mechanically less stable regions, whereas intramembranous bone formation takes place internal to the periosteum. Pro-inflammatory cytokines, members of the TGFß superfamily and angiogenic factors regulate the healing process, which is devided into four phases: (1) immediately after injury, inflammatory cytokines are produced that recruit MSCs into the hematoma; (2) these MSCs proliferate and differentiate into chondrocytes forming the so-called “soft callus”; (3) next, the chondrocyte matrix becomes mineralized and transforms into “woven bone”, a process that includes matrix resorption and the continuous recruitment of progenitor cells; (4) finally, during secondary bone formation, the instable woven bone is remodeled into strong lamellar bone (Ai-Aql et al. 2008; Fazzalari 2011).
BMPs and BMPRs during osteogenic differentiation
During the phases of bone fracture healing, BMPs display various expression patterns. BMP2 is upregulated immediately after injury and can be detected during the entire process (Fig. 3). BMP5 and BMP6 also increase during the earlier phases regulating MSC proliferation. GDF5 is expressed from day 7 to day 14 corresponding to the second phase of fracture healing regulating chondrogenesis. During the remodeling of cartilage into woven bone, BMP3, BMP4, BMP7 and BMP8 plus BMP2, BMP5 and BMP6 are upregulated (Ai-Aql et al. 2008), whereas the expression of most bone-healing-related factors is downregulated in non-unions (Niikura et al. 2006).

Endochondral repair of a bone fracture depicting early (top right) and late (bottom right) vascular BMP events. Following injury of bone with vessel rupture and bleeding, hypoxic conditions favor a local burst of BMP secretion by the accumulating white blood cells, bone marrow stromal cells and platelets. Neovascularization is primed by vasculogenesis forming a primitive vascular plexus via BMP-dependent recruitment and local differentiation of endothelial progenitor cells (a). BMP induces not only the differentiation of bone progenitor cells but also vascular endothelial growth factor (VEGF) expression and secretion forming a cooperatively acting BMP/VEGF gradient to be sensed by angiogenic sprouts of pre-existing vessels. Sprouting angiogenesis is induced by VEGF receptor 2 (VEGFR2) expression in a selected tip cell and is repressed in the stalks via Delta-like factor 4 (Dll4)/Notch activation of transcription factors Hey and Hes (Hey/Hes), also regulated by Smads (b). Fine tuners Gremlin and BMP-binding endothelial-cell-precursor-derived regulator (BMPER) balance the angiogenic response by binding BMPs regulating their endothelial bioavailability. BMP-dependent expression of matrix-metallo-proteases (MMPs) is important for the transition phase and for tissue remodeling with soft callus becoming hard callus (c). BMPs act on VEGF production originating from chondrocytes thereby recruiting vessels from the perichondrium, a procedure termed the angiogenic switch. This recruitment contributes to making soft callus become hard callus, since it triggers the resorption of cartilage and its replacement by bone through osteoclasts. Interestingly, pericytes lining the invading blood vessels might also promote ossification in an alternative mode, by providing additional bone progenitor niches from the periphery. Blood-vessel-forming cells have also been shown to dedifferentiate into multipotent progenitor cells in a BMP-dependent manner. At the same time, previously established new blood vessels need to mature via the coverage of vascular smooth muscle cells (VSMC), which also respond to BMPs by, for example, maintaining their phenotype. The final endothelial maturation goes hence with hard callus maturation characterized by reduced endothelial migration and proliferation, rather than quiescence induced by BMP9, which becomes delivered by the blood stream (d)
BMP2 is essential for the initiation of fracture healing. Mice with impaired Bmp2 expression in their limbs (Bmp2 c/c mice) show normal skeletal development but impaired fracture healing. Although other BMPs can compensate for the lack of BMP2 during bone development, none is able to substitute for the function of BMP2 during bone healing (Tsuji et al. 2006). During endochondral ossification, BMP2 regulates the recruitment of osteoprogenitor cells mainly from the periosteum and by the activation of differentiation in the chondrogenic and osteogenic lineages. Mice treated with rhBMP2 after fracture show enhanced chondrogenesis and an enlarged callus (Yu et al. 2010). By contrast, mice lacking BMP4 or BMP7 exhibit normal fracture healing (Tsuji et al. 2008, 2010). In addition to BMP2 and BMP7, which are used clinically, BMP6 and BMP9 have a high osteogenic potential as shown by the induced ALP activity, the expression of osteocalcin, and in vivo experiments (Kang et al. 2004; Luu et al. 2007).
The function of BMPs also depends on their associated receptors. BMPRIb-mediated signaling seems to be more important for osteogenic differentiation, whereas signaling through BMPRIa is more important for other differentiation processes. Truncated (tr)BMPRIb significantly reduces the expression of Runx2, ALP and Osteocalcin and inhibits matrix mineralization in 2T3 cells. Constitutively active (ca)BMPRIb induces ligand-independent Smad1 phosphorylation and matrix mineralization in those cells. In contrast, trBMPRIa blocks adipocyte differentiation, whereas caBMPRIa induces adipogenesis (Chen et al. 1998). The role of BMPRIb has also been investigated in vivo by the generation of transgenic mice referred to as tg(Col-2.3) expressing dominant-negative BMPRIb in osteoblasts. These mice exhibit impaired bone formation as shown by reduced bone mineralization, bone volume and bone formation rates, whereas osteoblast and osteoclast numbers are unchanged. In tg(Col-2.3) osteoblasts, BMP signaling is blocked and the cells do not produce a mineralized matrix (Zhao et al. 2002).
Osteogenic Smad and non-Smad signaling
The expression of Runx2, an early and essential transcription factor during osteogenic differentiation, is induced by the Smad pathway, as has been shown by dominant-negative Smad1, Smad5, or Smad4. Complex formation with activated Smad1/5 enhances the transcriptional activity of Runx2 but is not required for the osteogenic effects of Runx2 (Nishimura et al. 2002). Once expressed, Runx2 directly induces the expression of Osteocalcin, Osteopontin and CollagenI (Franceschi and Xiao 2003). Furthermore, Runx2 and Dlx5 stimulate ALP expression in an additive manner (Kim et al. 2004). Id1–4 are target genes of Smad signaling (Miyazawa et al. 2002). Several studies show that these transcription factors promote osteogenic differentiation by inhibition of the myoblastic differentiation process (Nojima et al. 2010). Id1/Id3 heterozygous knockout mice have decreased levels of late osteogenic markers such as Osteopontin and Osteocalcin and show reduced numbers of osteoblasts in combination with impaired mineralization but non-altered levels of Runx2, ALP and CollagenI. Proliferation of primary calvarial osteoblasts is clearly reduced in Id1/Id3-deficient mice; this is thought to be a reason for impaired bone formation (Maeda et al. 2004).
BMP-mediated activation of p38, ERK, JNK, or PI3K resembles the non-Smad signaling cascades during bone healing. Thereby, Smad and non-Smad signaling pathways synergistically act to activate and further enhance the function of important osteogenic transcription factors such as Dlx5, Runx2 and Osterix. To induce the expression of Dlx5, for example, active Smad complexes are required. To enhance the activity of this transcription factor, the Dlx5 protein is phosphorylated at Ser-34 and Ser-217 by activated p38 (Ulsamer et al. 2008). Concerning Runx2, the TAK1-MKK3/6-p38 MAPK pathway is required for further phosphorylation, even though Runx2 expression is also induced by Smad-dependent signaling. Runx2 modification by the TAK1-MKK pathways is found in Mkk3 −/− and Mkk6 +/− knockout mice, which show a decreased expression of Runx2 target genes such as ALP, Osteocalcin, Osterix, and CollagenI plus reduced ALP activity and matrix mineralization (Greenblatt et al. 2010). Moreover, Osterix is regulated by p38 as shown by the inhibition of the p38 pathway in primary mice osteoblasts; this inhibition results in the decreased expression of Osterix at both the mRNA and the protein levels (X. Wang et al. 2007). P38 phosphorylates Osterix to increase its transcriptional activity (Ortuno et al. 2010).
JNK is involved in late osteoblastic differentiation processes. Inhibition of the JNK pathway significantly reduces matrix mineralization in MC3T3-E1 cells and late osteogenic markers such as Osteocalcin and Bone sialoprotein are downregulated, whereas the expression of Runx2, Osterix and Osteopontin and the activity of ALP are not affected (Matsuguchi et al. 2009).
The role of the lipid kinase PI3K in osteogenic differentiation is not completely understood and the literature reports conflicting data on this topic. A positive role for PI3K in osteogenesis has been shown in mice with a double knockout of the PI3K target genes Akt1/Akt2. In addition to other defects, these mice show delayed bone ossification (Peng et al. 2003). The single knockout for Akt1 possesses shorter bones and impaired secondary ossification (Ulici et al. 2009). Furthermore, specific inhibition of PI3K blocks BMP2-induced osteogenesis as shown by the decreased expression levels of Dlx5, Runx2, Osterix and Osteocalcin and reduced ALP activity and mineralization in C3H10T1/2 cells (Mukherjee and Rotwein 2009). Constitutive active Akt positively regulates osteoblastic differentiation (Raucci et al. 2008). In contrast to the bone-stimulating functions, other groups have described a negative role for the PI3K/Akt pathway during osteoblastic differentiation. Inhibition of PI3K increases mineralization and the expression of Runx2 and Osterix in skeletal-muscle-derived stem cells from mice and also elevates ALP activity in C2C12 cells (Payne et al. 2010; Vinals et al. 2002).
In summary, the activation of both the Smad and the non-Smad pathways is obviously essential for fracture healing. However, fine-tuning and pathway crosstalk are important to enhance bone regeneration and have to be taken into account with resepct to clinical applications.
Crosstalk between endothelium and bone
Current investigations aim to uncover BMP-dependent mechanisms in the development and pathology of blood vessel formation, termed “vascular” BMP signaling. Even though not part of the musculoskeletal system, the contribution of the vasculature to repair and regeneration processes of especially bone is crucial. Inappropriate neo-vascularization during bone repair has been shown to be one cause for fibrous non-unions and delayed healing rates (Lu et al. 2007; Matuszewski and Mehta 2011; Geris et al. 2010). Abberant vessel formation during bone repair in mouse bone-defect models has revealed that healing rates are significantly decreased when vessels near the fracture site are diminished (Wallace et al. 1995; Fassbender et al. 2011). In the following paragraphs, we will describe the process of endochondral fracture healing and focus on the early and late “vascular BMP crosstalk” between the cell types that contribute to establishing a new vessel network. Vessel formation is crucial during early timepoints of bone regeneration and primes for a succesful healing outcome (Hankenson et al. 2011). New blood vessels that arise during the early phase of callus formation not only deliver nutrition and oxygen to the site of ossification but also provide important growth factors, immune-responsive cells and precursor-cells for tissue remodeling. Blood vessels transport waste products of necrotic tissue and deliver cells that negatively fine-tune osteogenesis, e.g., osteoclasts or chondroclasts. Most fractures include ruptures of micro-vessels or, in severe cases, damage and bleeding of torn periosteal vessels (Fig. 3a). Before the vessel closes, the bleeding leads to a local hemorrhage establishing a blood clot around the fracture gap (Kolar et al. 2010, 2011). Together with invading fibroblasts, this hematoma provides a first stabilizing structure and provides an ECM to which growth factors can bind and where cells can accumulate (Bruckner-Tuderman et al. 2010). White blood cells play a pivotal role here (Schmidt-Bleek et al. 2011; Lienau et al. 2009). They are the main source of cytokines inducing BMP expression in adjacent bone marrow stromal cells (Rifas 2006). Another consequence of this vascular disruption is the activation of platelets, which is characterized by growth factor release from granules (Gruber et al. 2002). BMP-containing platelet supernatants and platelet-rich plasma have been shown to induce the differentiation of bone precursor cells and enhance bone formation (Verrier et al. 2010; Chevallier et al. 2010; Anitua et al. 2004; Park et al. 2008). The identification of distinct BMP members that contribute at certain stages to human bone repair is difficult; however, most spatiotemporal expression profiles have been obtained by using the mouse or rat critical-size-defect model, which allows comments to be made on early and late BMPs in fracture repair. Evidence has been presented showing that the sequential expression of individual BMPs follows a certain sequence: BMP2-6 are described in various species as those BMPs appearing in the initial phase of bone healing, whereas BMP4 seems to be redundant to BMP2 with respect to its osteoinductive capacity (Tsuji et al. 2006, 2008; Bostrom et al. 1995; Yu et al. 2010). The established local hypoxic conditions have been found to induce BMP2 expression subsequently in osteoblasts; this is understood as a feed-forward mechanism (Tseng et al. 2010). However, others have described hypoxia to induce the expression of ALK1 and Smad1/5 in endothelial cells (ECs; Tian et al. 2010).
Vascular and angiogenic responses mediated by BMPs
How is blood vessel formation initiated and what is the role of BMPs in this process? To date, the two known modes of vessel formation, namely vasculogenesis and angiogenesis, have been identified as contributing to bone regeneration. De novo vessel formation by the recruitment and differentiation of endothelial progenitor cells (EPCs) is called vasculogenesis. BMP-dependent EPC recruitment from the periphery has been described in the context of tumor vascularization (Zaidi and Nixon 2007). Others have reported that the recruitment of EPCs contributes directly to bone healing by using labeled bone-marrow-derived EPCs (Matsumoto et al. 2008). BMP2 has been demonstrated to induce the chemotaxis of EPCs (Raida et al. 2006). Furthermore, BMP has been shown to induce VEGF expression in MSCs. A bidirectional signaling relationship between MSCs and ECs during the development of vascular structures has recently been reviewed elsewere (Kirkpatrick et al. 2011). VEGF, the most potent inducer of endothelial migration and proliferation, appears in distinct isoforms and splice variants with non-redundant functions during normal and aberrant angiogenesis and fracture repair (Patil et al. 2011; Beamer et al. 2010). VEGF-A has been observed to act synergistically with BMP4 to recruit MSCs and induce cartilage formation and remodeling during endochondral ossification. VEGF-A is a transcriptional target of BMP signaling in pre-osteoblastic cells (Deckers et al. 2002). Recent research has demonstrated in vivo the cooperative effects of BMP2 and VEGF on bone healing. Scaffold-hydrogel implants with sequential release kinetics for BMP2 and VEGF have revealed that an initial burst of VEGF in combination with locally sustained BMP2 significantly enhances ectopic bone formation and neo-vascularization in rats (Kempen et al. 2009). Bone marrow stromal cells stably transduced with vectors encoding both VEGF-A and BMP2 genes have been seeded on coral scaffolds implanted into rabbit critical-size-defect models (Xiao et al. 2011). Combined expression increases bone formation and angiogenesis compared with single treatment controls. As shown in vivo, VEGF improves, whereas soluble VEGF receptor (VEGFR) inhibits BMP2-induced bone formation and bone healing through the modulation of angiogenesis (Peng et al. 2005).
The initial local differentiation of EPCs forming a primitive vascular plexus is followed by the second mode of vessel formation termed angiogenesis. Angiogenesis thus relies on pre-existing vessels that further sprout and are pruned in response to chemotactic guidance cues such as VEGF. For this network-remodeling process, a single migrating tip cell becomes selected and activated within the vessel wall (Fig. 3b). It polarizes and responds to gradients of VEGF via VEGFR-associated pathways (Gerhardt 2008; Graupera et al. 2008; Nilsson et al. 2010). The contribution of BMP to this process is not entirely clear. The expression of VEGFR2, a high affinity receptor for VEGF-A, has been shown to increase gradually during callus formation (Reumann et al. 2010) suggesting that it correlates with increased vascularization. Miyazono and co-workers have shown that BMP4 treatment of microvascular ECs increases not only the expression levels of VEGFR2 but also the phosphorylation of VEGFR2 and angiopoetin receptor; treatment with BMP4 has direct effects on EC migration and proliferation in mouse embryonic stem-cell-derived ECs and human microvascular ECs (Suzuki et al. 2008). Recently, vascular BMP signaling has been demonstrated to regulate sprouting angiogenesis in zebrafish but only in axial veins independent of VEGF-A signals, whereas the classic VEGFR/VEGF-mediated mode of sprouting is found in the dorsal aorta (Wiley et al. 2011). To distinguish between VEGFR- and BMPR-mediated signaling, BMP pathway inhibitors such as Dorsomorphin are currently being used; however, we should take into consideration that, especially for kinases of the VEGFR family, these inhibitors display strong off-target effects (Vogt et al. 2011; Hao et al. 2010).
Pathways considered to transduce migratory BMP signals belong to the non-Smad signaling branch. In mouse myoblast cells, the PI3K and MAPK pathways have been shown to contribute to BMP-induced migration, regulating the activity of Rho GTPases and F-actin capping proteins, respectively (Gamell et al. 2008, 2011). Within the sprout, the single tip cell is followed by exclusively proliferating stalk cells, forming the lumen after fusion (Adams and Alitalo 2007). Both tip cells and stalk cells respond to VEGF-A, although by different means. During the migration of the tip cell, the stalk cell proliferates instead of migrating (Fig. 3b). The Delta-like factor 4 (Dll4)-Notch pathway has been described to play a leading role in this divergent response. VEGF induces the expression of Dll4 in EC tips. A negative feedback loop by Dll4 acts on Notch receptor activation in adjacent stalk cells downregulating VEGFR expression via the transcriptional regulators Hey and Hes (Taylor et al. 2002; Adams and Eichmann 2010; Suchting et al. 2007). The genetic disruption of Notch in mice causes an increase in the number of tip cells, resulting in enhanced vascular sprouting (Hellstrom et al. 2007). BMP2 has been shown not only to transactivate Notch signaling but also directly to influence Notch pathway proteins. Hes and Hes-related proteins such as Hey1 are induced by BMP stimulation in various cells, including osteoprogenitor cells and ECs (Miyazono et al. 2005). BMPs have been found to upregulate Notch ligand JAG1 directly via Smad1/5 signaling in ECs (Morikawa et al. 2011). The activation of Hey and Hes genes by Notch/Dll4 and BMP signaling have been proposed to act in a cooperative manner, since Smad1 and NIC, the shedded intracellular domain of Notch, form a complex (Takizawa et al. 2003). In ECs, Hey1 has been demonstrated to be synergistically induced by both the BMP and Notch signaling pathways (Itoh et al. 2004). In neuronal epithelial cells, BMP2 acts as an inhibitor of neurogenesis, as is partly described by the BMP2-induced phosphorylation of Smad1/5/8 and binding to the consensus sequence in the Hes-5 promoter (Nakashima et al. 2001). BMP-Notch crosstalk has also been described for osteoblasts (de Jong et al. 2004; Hilton et al. 2008), identifying Notch signaling as inhibiting osteogenic differentiation by diminishing Runx2 transcriptional activity via Hes (Hilton et al. 2008; Zamurovic et al. 2004). An example of the way that functional crosstalk between the VEGF, TGFβ and Notch signaling pathways can pattern the vasculature has previously been presented (Holderfield and Hughes 2008) and therefore, a related mechanism probably also uses BMP-specific signaling.
Taking together, vascular BMP signaling in the initial phase of bone healing triggers VEGF expression and secretion in mesenchymal bone progenitor cells and bone marrow stromal cells, establishing a paracrine VEGF pool that can be sensed by circulating EPCs or endothelial tip cells of proximal vessels. These respond by inducing vasculogenesis or angiogenesis, respectively. In addition to this paracrine action, BMPs act in a direct fashion on EC physiology, by using the BMP pathway and its receptors to induce signaling in cooperation with VEGFs and also the Notch pathway mediating EC proliferation and migration. However, the molecular mechansisms underlying the latter have not yet been entirely deciphered.
Fine tuners of vascular BMP signaling
Gremlin and BMPER have recently been discovered to act as fine tuners of vascular BMP signaling. Gremlin acts as a BMP inhibitor by forming heteromers sequestering BMP2, BMP4 and BMP7 from BMPR interaction (Fig. 3c; Claesson-Welsh 2010). In addition, gremlin is able to bind VEGFR2 independently of complex formation with BMPs, initiating VEGFR2 signaling and thereby mimicking the action of VEGFs (Mitola et al. 2010). Interstingly, the simultaneous delivery of gremlin and BMP4 to the chick chorioallantoic membrane (CAM assay) triggers a more potent angiogenic response when compared with gremlin alone (Stabile et al. 2007) suggesting that a BMP/gremlin complex acts cooperatively to induce VEGFR2-dependent angiogenesis. BMPER, on the other hand, is an ECM protein secreted by ECs (Moser and Patterson 2005; Moser et al. 2003). BMPER binds directly to BMPs and has a dose-dependent stimulating effect on BMP-induced EC sprouting and migration (Heinke et al. 2008). BMPER dynamically switches from an activator to an inhibitor of BMP4 signaling in a dose-dependent fashion and is thus postulated to exert pro- and anti-BMP activities (Kelley et al. 2009). In ECs, BMP6 has been shown to activate Smad1/5/8 phosphorylation leading to the transcriptional activation of Id1, whereas the ectopic expression of Id1 leads to EC migration and tube formation (Valdimarsdottir et al. 2002). BMP6-induced tube formation is blocked by the knockdown of Id1. The role of Ids in vascular BMP signaling is not entirely clear; however, they are proposed to regulate matrix-metallo-proteases (MMPs) and integrins as has also been described for other cell types (Fig. 3c; Alani et al. 2004; Song et al. 2011; Su et al. 2010; Darby et al. 2008; Ronziere et al. 2005). During fracture healing, the fine tuning role of MMPs is to remodel the ECM (Kasper et al. 2007; Behonick et al. 2007; Lieu et al. 2011), regulate matrix stiffness and release pools of matrix-bound growth factors and other vascular mediators (Medley et al. 2004; Lehmann et al. 2005; Ebrahem et al. 2010).
Vascular maturation and late steps of endochondral ossification
BMPs are proposed to have distinct functions in EC activation versus maturation (David et al. 2009). BMP9 and BMP10 bind to the high affinity receptor ALK1 and co-receptor endoglin, both primarily expressed in ECs, even though BMP10 expression is restricted to the heart (David et al. 2007a; Neuhaus et al. 1999). Alk1 −/− mice display early embryonic lethality because of vascular abnormalities in both the yolk sac and embryo, similar to the phenotype of endoglin disruption (Oh et al. 2000; Urness et al. 2000; Li et al. 1999; Arthur et al. 2000). ALK1 directly phosphorylates endoglin, resulting in the inhibition of EC proliferation (Koleva et al. 2006). Interference with ALK1 activation in human umbilical cord vascular endothelial cells (HUVEC) has revealed the failure of endothelial cord formation in matrigel assay and reduced VEGF- and BMP10-mediated vessel formation in the CAM assay. Amongst all BMPs, BMP9 represents one of the rare biologically active forms circulating in the blood stream and has been described as inhibiting EC migration and proliferation suggesting that it is a vascular quiescence factor (David et al. 2007b; Scharpfenecker et al. 2007; K. Wang et al. 2011). However, controversial results in ECs of a different origin have also demonstrated that BMP9 is able to activate ECs (Suzuki et al. 2010). Of special interest is that BMP9 is resistant to noggin inhibition and therefore underlies a different type of feedback regulation in comparison to BMPs activating ECs (Seemann et al. 2009). Taken together, research focusing on BMP9 suggests that its vascular role is highly EC-type specific and that its regulatory mechanisms might largely differ from those known for other BMP family members. Since BMP9 cycles in the blood stream, it exerts ubiquitous bioavailability and is likely to be the first BMP sensed by ECs from the inside of a freshly formed blood transfusing lumen.
In addition to BMP9, other BMPs have been proposed to be involved in the maturing processes of bone and the surrounding endothelium by mechanisms inducing cell survival, quiescence and phenotype maintenance. BMP2 is present throughout bone repair and thus its contribution to bone healing and endothelial crosstalk at later stages differs from its early roles and it appears to act in concert with exclusively late-expressed BMPs. During these later stages, periosteal cells proximal to the fracture gap develop into chondroblasts and periosteal cells distal to the fracture gap develop into osteoblasts. The so-formed soft callus undergoes endochondral (and intramembranous) ossification. Here, the avascular cartilage undergoes neo-vascularization followed by the recruitment of chondroclasts that contribute to the remodeling and substitution of the cartilage with lamellar bone via mineralization of the collagen matrix. In this process, the neo-vascularization of cartilage is understood as a switch triggering cartilage removal and substitution by bone. During this time, BMP3, BMP4, BMP7 and BMP8 display peaks of expression during the repair of mouse tibias (day 14 to day 21; Cho et al. 2002). At this time point, the resorption of calcified cartilage is most active. BMP2 has been shown to up-regulate VEGF-A, -B, and -C synthesis and VEGFR2 expression in chondrocytes (Bluteau et al. 2007). VEGF-A produced by hypertrophic chondrocytes induces osteoclastogenesis in the perichondrium and stimulates the migration of osteoclasts into hypertrophic regions (Henriksen et al. 2003; Patil et al. 2011; Higashikawa et al. 2006). Hypertrophic chondrocytes, which respond to BMP2 by the secretion of VEGFs, act again not only in an autocrine but also in a paracrine manner. The autocrine process is needed for chondrocyte survival (Maes et al. 2004; Zelzer et al. 2004; Zelzer and Olsen 2005), whereas paracrine processes might further recruit ECs needed for the neovascularization of the hypertrophic region. BMP-induced VEGF signaling in chondrocytes is therefore crucial not only for endochondral ossification but also for the maturation of the endothelium in the distal callus areas.
Whereas the distal parts of the fracture are already mineralized and vascularized, the proximal parts are still soft and avascular. The blood vessels that have become established at the periosteum during the initial phase of repair need to mature in order to achieve full functionality; however, neovascularization of the remodeling region also needs to be promoted as mentioned above (Fig. 3d). EC maturation is characterized by quiescence and phenotype maintenance. Recent research on the plasticity of ECs has surprisingly revealed that BMP4 or TGFβ2 treatment is sufficient to dedifferentiate mature ECs into a multipotent MSC-like phenotype, providing first insights into the potential of ECs as bone-forming cells via BMP-dependent endothelial to mesenchymal transition (Medici et al. 2010; Horwitz 2010). Endothelial maturation and functionality is also achieved by the additional recruitment and alignment of cells covering the lumen from outside (Fig. 3d). Perivascular cells lining the endothelial vessel wall are summarily referred to as pericytes and include vascular smooth muscle cells (Ribatti et al. 2011; Caplan 2009) important for endothelial maturation and survival via bidirectional signaling (Gaengel et al. 2009; Hellberg et al. 2010; Franco et al. 2011). BMP2 and BMP7 inhibit the proliferation of serum-stimulated pulmonary artery SMCs (Morrell et al. 2001). BMP4 has been shown to inhibit migration but does not affect SMC proliferation (Corriere et al. 2008). Recent studies by various laboratories have identified pericytes as having mesenchymal bone-progenitor-like properties and a huge regenerative potential, making them an alternative source of bone precursors that can influence ossification from the distal part of an injury (Maes et al. 2010; Corselli et al. 2010; Crisan et al. 2011; Chen et al. 2009). The bone-precursor-providing niche could therefore not only be found in bone marrow but might also be provided by blood vessels. Bone neo-vascularization might then promote repair in a secondary mode, alternatively delivering, from the periphery, MSC niches that help to initiate intramembranous ossification. The role of BMPs in this process is not entirely clear since, in addition to their differentiation potential on precursor cells, they have been observed to maintain SMC phenotypes (Dorai et al. 2000). SMCs covering the endothelium mediate EC cell contacts, thereby being crucial players influencing structural integrity. Cytokines secreted by SMCs participate in the regulation of EC adhesion molecule expression, such as vascular cell adhesion proteins (VCAMs). Interestingly, Smad1 null mice die at embryonic day 9.5 and display disorganized vessel formation and a lack of VCAM1 expression (Lechleider et al. 2001). The dysregulation of BMP-SMC signaling has also been implicated in vascular calcification, hypertrophy and artherosclerotic plaques (Nakagawa et al. 2010; Hansmann et al. 2008; Schluesener and Meyermann 1995).
In summary, the effects of BMPs in blood vessel formation during bone repair are diverse, exerting early and late vascular activity. A variety of cell types respond to BMPs during vascularization events and tightly crosstalk to each other by modes of paracrine and autocrine endothelial activation and maturation. Clearly, the spatiotemporal expression patterns of BMP subtypes and the respective fine tuners are important for stepwise and controlled ossification.
BMP signaling in the central and peripheral nervous system
During vertebrate development the branching of blood vessels appears similar to the branching of nerves and researchers are searching for shared regulatory proteins (Autiero et al. 2005). BMP signaling has been shown also to govern structural and physiological functions in neurons (Liu et al. 2003; Liu and Niswander 2005; Hall and Miller 2004). Early research on BMP signaling in neurons was associated with investigations of dorso-ventral axis formation in Drosophila. This research revealed that BMPs (BMP2/4 homolog = Decapentaplegic [Dpp]) suppress neurogenesis, thus defining non-neural ectoderm, whereas the inhibition of BMP signaling by antagonistic gradients of chordins gives rise to neural ectoderm (Holley et al. 1995; Biehs et al. 1996). The neural ectoderm forms, during later developmental stages, the central and peripheral nervous systems (CNS/PNS). Although first shown in Drosophila, this mechanism is conserved and is also found in vertebrates. In contrast to the establishment of neural tissue, BMPs have been shown to act positively on neuronal precursor cell differentiation and proliferation, for example, during embryonic spinal tube formation, together with Wnts (Chesnutt et al. 2004). BMPs are also important for astrocyte formation (D’Alessandro et al. 1994), the guidance of axonal growth cones (Augsburger et al. 1999), glial cell maturation (See et al. 2007) and finally the coordinated assembly of the synapse. Past research has focused on BMPs and their function as potential neurotrophic and neuroprotective factors to tackle traumatic CNS and PNS injury (Sabo et al. 2009; Helm et al. 2000). BMP7 promotes, for example, the survival of glial cells after acute traumatic injury in the rat spinal cord injury model. BMP7 mRNA is upregulated only in glial cells post-injury, suggesting that BMP7 regulates the differentiation of glial cells from neural progenitors and induces gliosis after CNS trauma (Setoguchi et al. 2001). Other investigations deal with the inhibition of spinal cord recovery after injury mediated by BMPs. BMP2 and BMP4 have been shown to exert elevated expression in oligodendrocytes and astrocytes around sites of spinal cord injury, whereas the administration of Noggin at these sites leads to enhanced locomotor activity and significant re-growth of the spinal cord (Matsuura et al. 2008). Similar to their vascular roles, these observations might again rely on spatiotemporal expression patterns of both ligands and receptors, especially under pathophysiological conditions. GDF15 is a distant member of the TGFβ superfamily and is described as a novel trophic factor for motor and sensory neurons. It is expressed in Schwann cells and promotes the survival of cultured motor, sensory and sympathetic neurons (Strelau et al. 2009). Elevated GDF15 expression is a hallmark of many tissue injuries, including those in the kidney, liver and lung. GDF15 mRNA is upregulated in lesioned neurons (Strelau et al. 2003) and leasoned brain (Schindowski et al. 2011), whereas it is barely detectable in unlesioned CNS and intact peripheral nerves (Strelau et al. 2000). GDF15-deficient mice display progressive loss of spinal, facial and trigeminal motoneurons accompanied by significant impairment of motor skills (Strelau et al. 2009). The signaling mechanisms underlying the neuroprotective function of GDF15 involve the non-Smad cascades such as PI3K/Akt and Erk (Subramaniam et al. 2003).
BMP crosstalk at the neuromuscular junction
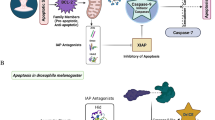
Interestingly, BMPs have been found to play an important role in proper gross and fine motor skills. BMPs mediate the formation of the neuromuscular junction (NMJ) in which motor neurons transmit signals from the PNS toward effector muscles (Fig. 4). The Drosophila BMP5-8 homolog glass bottom boat (Gbb) has been shown to regulate the growth of synapses and their maturation, neurotransmission and homeostasis (McCabe et al. 2003; Baines 2004; Goold and Davis 2007). BMPs and their role in NMJ signaling are of special interest, since aberrant NMJ signaling is involved in a multitude of neurodegenerative disorders such as spinal muscular atrophy, multiple sclerosis and Huntington’s disease (Salinas et al. 2008; Bayat et al. 2011). Fly genetics has shown that Gbb secreted from the muscle is required for the transcriptional regulation of the guanosine-exchange-factor Trio in pre-synaptic cells, thereby providing another example of paracrine crosstalk. In the pre-synapse, Trio in turn activates small GTPases regulating cytoskeletal rearrangements necessary for subsequent controlled synaptic outgrowth (Ball et al. 2010; Fuentes-Medel and Budnik 2010). The pre-synaptic BMP responsiveness is fine tuned and balanced by endocytic events of the Drosophila BMPRI (Drosophila homolog: thickveins [Tkv]; O’Connor-Giles et al. 2008). Dynamin and Dap160/intersectin have been demonstrated to inhibit muscle-originated BMP signaling by promoting receptor endocytosis at the pre-synapse. Interference with components of this endocytic machinery lead to excessive NMJ growth. On the other hand, the Dawdle/Baboo pathway (Drosophila homolog of activin ligand/ACTRI: Daw/Babo) acts upstream of muscular Gbb expression emphasizing an activin/BMP signaling loop at the NMJ (Ellis et al. 2010; Bayat et al. 2011). The Smad analog in Drosophila is termed mothers against decapentaplegic (Mad). In strong accordance with the vertebrate pathway, Gbb or Dpp mediate the phosphorylation of Mad modulating the expression of Drosophila target genes. The downstream signaling events necessary to mediate BMP-induced NMJ formation are not well characterized but probably also include the Smad-independent activation of LIMK1 involved in cytoskeletal rearrangements and migration (Foletta et al. 2003). BMP7 gradients have been shown to steer nerve growth cones of Xenopus laevis in a LIMK1-dependent fashion (Wen et al. 2007). In Drosophila and chicken, LIMK1 has been implicated in the mechanisms of axonal outgrowth through the LIMK-ADF/cofilin pathway (Arber et al. 1998; Ohashi et al. 2000; Meberg 2000; Endo et al. 2007). LIMK1 interacts directly with the cytoplasmic tail-region of BMPRII (Foletta et al. 2003) and was originally isolated in screens for novel kinases expressed in the CNS/PNS (Kang et al. 2004; Cheng and Robertson 1995; Proschel et al. 1995). Wishful thinking (Wit) is the Drosophila homolog of BMPRII. In Wit mutant larvae, the size of the NMJs is greatly reduced relative to the size of the muscles. NMJ phenotypes can be rescued by the transgenic expression of Wit only in motoneurons (Aberle et al. 2002).

BMP crosstalk at the Drosophila neuromuscular junction (NMJ). Correct assembly at the NMJ relies on Glass bottom boat (Gbb) signaling from the muscle toward the motor neuron. Activin induces BMP/Gbb secretion by muscle cells; this in turn activates complex of BMP type I receptors Thickveins (Tkv), Saxophone (Sax) and type II receptor Wishful thinking (Wit) at the pre-synapse. By Dynamin/Intersectin-dependent endocytosis, the receptor complex becomes endocytosed and phosphorylates Mothers against Decaplentaplegic (Mad) releasing the receptor complex to translocate into the nucleus. There, Mads regulate the expression of Triple functional domain protein (Trio), an exchange factor important for Rho GTPase activity at the site of F-actin formation and cytoskeletal rearrangements. LIMK has also been shown to be involved in neuronal guidance and F-actin assembly. (LIMK) interacts and becomes activated by Wit
In summary, the establishment and amount of NMJs are tightly connected to functional BMP signaling in a paracrine manner, originating from the muscle but acting on the pre-synapse. BMP signaling at the synapse includes the activation of small GTPases, LIMK1 and Smads leading to guidance and growth toward muscles. Further identification of BMP-regulated key players in musculoskeletal innervation might therefore be a good strategy not only to address the urgent need for successful CNS/PNS trauma therapy but also to treat genetic diseases involving dysregulated signaling at the NMJ.
BMP signaling in cartilage repair
In contrast to bone and muscle, the tissue crosstalk of articular cartilage is limited because of poor vascularization and innervation and therefore, cartilage has a relatively low regeneration capacity after injury. Current treatments of osteoarthritis (OA), a chronic degenerative joint disorder of worldwide occurrence are arthroplasty and autologous chondrocyte transplantations. However, the recovery process is insufficient because of the development of fibrocartilage in large areas of the defect (O’Driscoll 1998). The gold standard for the treatment of small lesions below 4 cm2 is microfracturing, which induces and stimulates bone-cartilage interplay (Steinwachs et al. 2008). Microfracturing is defined by the penetration of the subchondral bone plate triggering bone marrow stimulation, migration, proliferation and the chondrogenic differentiation of MSCs invading the injured area from the bone marrow. In vitro approaches of chondrogenic differentiation are usually performed on bone-marrow-derived stem cells (Kuroda et al. 2007; Wakitani et al. 2004) or muscle-derived stem cells (Adachi et al. 2002). Transcription factors known to be involved in early chondrogenic processes and the control of GDF5 expression are members of the Hox and Sox gene families. Mutations of hoxa and hoxd genes cause fusions of the carpal joints (Favier et al. 1996). GDF5 (BMP14, CDMP1) is one of the most prominent players during chondrogenic differentiation. In the developing chick limb, overexpression of GDF5 increases the size of chondrogenic skeletal elements through the stimulation of cell proliferation (Francis-West et al. 1999). The GDF5 deficiency in the brachypod mouse results in skeletal malformations caused by defects in the formation of the mesenchymal condensations preceeding chondrogenesis and bone fusion (Gruneberg and Lee 1973; Storm et al. 1994). In humans, brachydactylies (BDs) and synostosis are caused by GDF5 point mutations (Schwaerzer GK et al. 2011; Seemann et al. 2009; Everman et al. 2002). BDs are characterized by the shortening of the digits because of malfunctions during patterning, condensation and differentiation of the phalanges or metacarpals, caused by a disrupted GDF5 signaling pathway. Several types of BD and the corresponding mutations within the genes encoding GDF5, BMPRIb and Ror2 have been described as targeting both the Smad and non-Smad signaling pathways (Polinkovsky et al. 1997; Lehmann et al. 2003; Sammar et al. 2004). BMPRIb is the type I receptor necessary for cartilage development (Ashique et al. 2002). Expression of the dominant negative BMPRIb but not BMPRIa has been shown to block embryonic chondrogenesis (Oshin and Stewart 2007). Postnatally, BMPRIa is expressed in prehypertrophic chondrocytes indicating functions during cartilage homeostasis (Rountree et al. 2004; Zou et al. 1997). The mechanisms described above are important steps during embryonic cartilage development. An analysis has been undertaken to determine whether they are also essential for adult cartilage repair. Indeed, BMP2 expression in healthy cartilage is low and increases in areas around cartilage lesions in OA (Blaney Davidson et al. 2006) and after mechanical injury (Dell’Accio et al. 2006). BMP2 exposure to healthy and damaged cartilage has been shown to increase proteoglycan synthesis and elevate the degradation of aggrecan mediated by MMPs, indicating BMP2 action in the replacement of damaged cartilage matrix molecules (Blaney Davidson et al. 2007). Overexpression of BMP4 in MSCs encapsulated in three-dimensional scaffolds leads to the expression of chondrogenic markers such as chondroitin-4-sulfate and chondroitin−6-sulfate and type II collagen (Steinert et al. 2003). Retroviral transduction of BMP4 into muscle-derived stem cells leads to the development of a chondrocytic phenotype (Kuroda et al. 2006). In addition to stimulatory effects on chondrocytes to express cartilage-specific extracellular proteins, BMP7 seems to have an anabolic effect on chondrocytes by promoting cell survival, proliferation and the regulation of anabolic pathways such as the IGF pathway (Chubinskaya et al. 2007).
Concluding remarks
During repair processes, the musculoskeletal system acts in concert with adjacent tissues through paracrine and autocrine signaling; this is thought to recapitulate major aspects of development. Bone is a well-connected tissue embedded into innervated muscle and tendon and is additionally supplied by blood vessels. Relative to the more poorly connected cartilage, muscle and bone display good regenerative potential. A variety of soluble growth factors mediate repair. In this review, our aim has been to highlight BMPs and their leading role in repair by referring to the recent literature describing both in-vitro- and in-vivo-based research. Apart from their classical morphogenic function inducing the differentiation of bone precursor cells, BMPs exert cellular responses ranging from migration to homeostasis and including the proliferation and apoptosis of cells of diverse origins. Interestingly, in highly connected tissues, a variety of BMPs appear in a spatiotemporal fashion during repair, starting with early detectable BMPs and continuing with inflammatory processes up to the later stages in which BMPs are involved in remodeling and maturation. Nevertheless, to date, only a few distinct BMP members have been discovered to be involved in the repair of more poorly connected tissues. Individual BMPs exhibit high affinity to certain BMPRs and also to co-receptors, ECM and fine tuning antagonists in a tissue-specific manner. Our current goal is to understand these differential affinities and the tissue-specific effects that have a major impact on the usage of recombinant BMPs in the clinic. Efforts in regenerative medicine are directed toward the use of BMPs alone or in combination with other growth factors to enhance repair. However, there is a substantial need to potentiate the effects of these factors further and to individualize treatments for a certain patient or injury situation. This includes the design of recombinant BMPs with altered affinities to receptors and antagonists and research into physiological release kinetics, implant-coating strategies, the use of a variety of biomaterials and delivery methods for gene therapy.
We have emphasized here the crosstalk potential of BMPs acting not only on the target tissue of repair but also on the connected tissues located in close proximity. In our view, the current task is to merge in vitro data at the subcellular level with animal models in order to gain insights into physiological repair processes at spatiotemporal resolution. To bridge the gap between these research methods, the further development of techniques mimicking nature will be needed. Current research on co-culture systems, bioreactors and advanced imaging methods will additionally allow us to understand the way that individual BMPs crosstalk to orchestrate specific tissue repair.
References
Aberle H, Haghighi AP, Fetter RD, McCabe BD, Magalhaes TR, Goodman CS (2002) Wishful thinking encodes a BMP type II receptor that regulates synaptic growth in Drosophila. Neuron 33:545–558
Adachi N, Sato K, Usas A, Fu FH, Ochi M, Han CW, Niyibizi C, Huard J (2002) Muscle derived, cell based ex vivo gene therapy for treatment of full thickness articular cartilage defects. J Rheumatol 29:1920–1930
Adams RH, Alitalo K (2007) Molecular regulation of angiogenesis and lymphangiogenesis. Nat Rev Mol Cell Biol 8:464–478
Adams RH, Eichmann A (2010) Axon guidance molecules in vascular patterning. Cold Spring Harb Perspect Biol 2:a001875
Ai-Aql ZS, Alagl AS, Graves DT, Gerstenfeld LC, Einhorn TA (2008) Molecular mechanisms controlling bone formation during fracture healing and distraction osteogenesis. J Dent Res 87:107–118
Alani RM, Silverthorn CF, Orosz K (2004) Tumor angiogenesis in mice and men. Cancer Biol Ther 3:498–500
Allouh MZ, Yablonka-Reuveni Z, Rosser BW (2008) Pax7 reveals a greater frequency and concentration of satellite cells at the ends of growing skeletal muscle fibers. J Histochem Cytochem 56:77–87
Amthor H, Christ B, Weil M, Patel K (1998) The importance of timing differentiation during limb muscle development. Curr Biol 8:642–652
Anitua E, Andia I, Ardanza B, Nurden P, Nurden AT (2004) Autologous platelets as a source of proteins for healing and tissue regeneration. Thromb Haemost 91:4–15
Arber S, Barbayannis FA, Hanser H, Schneider C, Stanyon CA, Bernard O, Caroni P (1998) Regulation of actin dynamics through phosphorylation of cofilin by LIM-kinase. Nature 393:805–809
Arthur HM, Ure J, Smith AJ, Renforth G, Wilson DI, Torsney E, Charlton R, Parums DV, Jowett T, Marchuk DA, Burn J, Diamond AG (2000) Endoglin, an ancillary TGFbeta receptor, is required for extraembryonic angiogenesis and plays a key role in heart development. Dev Biol 217:42–53
Ashique AM, Fu K, Richman JM (2002) Signalling via type IA and type IB bone morphogenetic protein receptors (BMPR) regulates intramembranous bone formation, chondrogenesis and feather formation in the chicken embryo. Int J Dev Biol 46:243–253
Augsburger A, Schuchardt A, Hoskins S, Dodd J, Butler S (1999) BMPs as mediators of roof plate repulsion of commissural neurons. Neuron 24:127–141
Autiero M, De Smet F, Claes F, Carmeliet P (2005) Role of neural guidance signals in blood vessel navigation. Cardiovasc Res 65:629–638
Baines RA (2004) Synaptic strengthening mediated by bone morphogenetic protein-dependent retrograde signaling in the Drosophila CNS. J Neurosci 24:6904–6911
Ball RW, Warren-Paquin M, Tsurudome K, Liao EH, Elazzouzi F, Cavanagh C, An BS, Wang TT, White JH, Haghighi AP (2010) Retrograde BMP signaling controls synaptic growth at the NMJ by regulating trio expression in motor neurons. Neuron 66:536–549
Barbara NP, Wrana JL, Letarte M (1999) Endoglin is an accessory protein that interacts with the signaling receptor complex of multiple members of the transforming growth factor-beta superfamily. J Biol Chem 274:584–594
Bayat V, Jaiswal M, Bellen HJ (2011) The BMP signaling pathway at the Drosophila neuromuscular junction and its links to neurodegenerative diseases. Curr Opin Neurobiol 21:182–188
Beamer B, Hettrich C, Lane J (2010) Vascular endothelial growth factor: an essential component of angiogenesis and fracture healing. HSS J 6:85–94
Behonick DJ, Xing Z, Lieu S, Buckley JM, Lotz JC, Marcucio RS, Werb Z, Miclau T, Colnot C (2007) Role of matrix metalloproteinase 13 in both endochondral and intramembranous ossification during skeletal regeneration. PLoS One 2:e1150
Berasi SP, Varadarajan U, Archambault J, Cain M, Souza TA, Abouzeid A, Li J, Brown CT, Dorner AJ, Seeherman J, Jelinsky SA (2011) Divergent activities of osteogenic BMP2, and tenogenic BMP12 and BMP13 independent of receptor binding affinities. Growth Factors 29:128–139
Biehs B, Francois V, Bier E (1996) The Drosophila short gastrulation gene prevents Dpp from autoactivating and suppressing neurogenesis in the neuroectoderm. Genes Dev 10:2922–2934
Birchmeier C, Brohmann H (2000) Genes that control the development of migrating muscle precursor cells. Curr Opin Cell Biol 12:725–730
Blaney Davidson EN, Vitters EL, Kraan PM van der, Berg WB van den (2006) Expression of transforming growth factor-beta (TGFbeta) and the TGFbeta signalling molecule SMAD-2P in spontaneous and instability-induced osteoarthritis: role in cartilage degradation, chondrogenesis and osteophyte formation. Ann Rheum Dis 65:1414–1421
Blaney Davidson EN, Vitters EL, Lent PL van, Loo FA van de, Berg WB van den, Kraan PM van der (2007) Elevated extracellular matrix production and degradation upon bone morphogenetic protein-2 (BMP-2) stimulation point toward a role for BMP-2 in cartilage repair and remodeling. Arthritis Res Ther 9:R102
Blitz E, Viukov S, Sharir A, Shwartz Y, Galloway JL, Pryce BA, Johnson RL, Tabin CJ, Schweitzer R, Zelzer E (2009) Bone ridge patterning during musculoskeletal assembly is mediated through SCX regulation of Bmp4 at the tendon-skeleton junction. Dev Cell 17:861–873
Bluteau G, Julien M, Magne D, Mallein-Gerin F, Weiss P, Daculsi G, Guicheux J (2007) VEGF and VEGF receptors are differentially expressed in chondrocytes. Bone 40:568–576
Bostrom MP, Lane JM, Berberian WS, Missri AA, Tomin E, Weiland A, Doty SB, Glaser D, Rosen VM (1995) Immunolocalization and expression of bone morphogenetic proteins 2 and 4 in fracture healing. J Orthop Res 13:357–367
Brohmann H, Jagla K, Birchmeier C (2000) The role of Lbx1 in migration of muscle precursor cells. Development 127:437–445
Bruckner-Tuderman L, Mark K von der, Pihlajaniemi T, Unsicker K (2010) Cell interactions with the extracellular matrix. Cell Tissue Res 339:1–5
Cain RJ, Ridley AJ (2009) Phosphoinositide 3-kinases in cell migration. Biol Cell 101:13–29
Caplan AI (2009) Why are MSCs therapeutic? New data: new insight. J Pathol 217:318–324
Carlisle E, Fischgrund JS (2005) Bone morphogenetic proteins for spinal fusion. Spine J 5:240S–249S
Carlson BM (2003) Muscle regeneration in amphibians and mammals: passing the torch. Dev Dyn 226:167–181
Chen YG, Massague J (1999) Smad1 recognition and activation by the ALK1 group of transforming growth factor-beta family receptors. J Biol Chem 274:3672–3677
Chen D, Ji X, Harris MA, Feng JQ, Karsenty G, Celeste AJ, Rosen V, Mundy GR, Harris SE (1998) Differential roles for bone morphogenetic protein (BMP) receptor type IB and IA in differentiation and specification of mesenchymal precursor cells to osteoblast and adipocyte lineages. J Cell Biol 142:295–305
Chen CW, Montelatici E, Crisan M, Corselli M, Huard J, Lazzari L, Peault B (2009) Perivascular multi-lineage progenitor cells in human organs: regenerative units, cytokine sources or both? Cytokine Growth Factor Rev 20:429–434
Cheng AK, Robertson EJ (1995) The murine LIM-kinase gene (limk) encodes a novel serine threonine kinase expressed predominantly in trophoblast giant cells and the developing nervous system. Mech Dev 52:187–197
Chesnutt C, Burrus LW, Brown AM, Niswander L (2004) Coordinate regulation of neural tube patterning and proliferation by TGFbeta and WNT activity. Dev Biol 274:334–347
Chevallier N, Anagnostou F, Zilber S, Bodivit G, Maurin S, Barrault A, Bierling P, Hernigou P, Layrolle P, Rouard H (2010) Osteoblastic differentiation of human mesenchymal stem cells with platelet lysate. Biomaterials 31:270–278
Chhabra A, Tsou D, Clark RT, Gaschen V, Hunziker EB, Mikic B (2003) GDF-5 deficiency in mice delays Achilles tendon healing. J Orthop Res 21:826–835
Cho TJ, Gerstenfeld LC, Einhorn TA (2002) Differential temporal expression of members of the transforming growth factor beta superfamily during murine fracture healing. J Bone Miner Res 17:513–520
Chubinskaya S, Hurtig M, Rueger DC (2007) OP-1/BMP-7 in cartilage repair. Int Orthop 31:773–781
Claesson-Welsh L (2010) Gremlin: vexing VEGF receptor agonist. Blood 116:3386–3387
Clark RT, Johnson TL, Schalet BJ, Davis L, Gaschen V, Hunziker EB, Oldberg A, Mikic B (2001) GDF-5 deficiency in mice leads to disruption of tail tendon form and function. Connect Tissue Res 42:175–186
Clever JL, Sakai Y, Wang RA, Schneider DB (2010) Inefficient skeletal muscle repair in inhibitor of differentiation knockout mice suggests a crucial role for BMP signaling during adult muscle regeneration. Am J Physiol Cell Physiol 298:C1087–C1099
Corriere MA, Rogers CM, Eliason JL, Faulk J, Kume T, Hogan BL, Guzman RJ (2008) Endothelial Bmp4 is induced during arterial remodeling: effects on smooth muscle cell migration and proliferation. J Surg Res 145:142–149
Corselli M, Chen CW, Crisan M, Lazzari L, Peault B (2010) Perivascular ancestors of adult multipotent stem cells. Arterioscler Thromb Vasc Biol 30:1104–1109
Crisan M, Corselli M, Chen CW, Peault B (2011) Multilineage stem cells in the adult: a perivascular legacy? Organogenesis 7:101–104
Dahlgren LA, Mohammed HO, Nixon AJ (2005) Temporal expression of growth factors and matrix molecules in healing tendon lesions. J Orthop Res 23:84–92
D’Alessandro JS, Yetz-Aldape J, Wang EA (1994) Bone morphogenetic proteins induce differentiation in astrocyte lineage cells. Growth Factors 11:53–69
Darby S, Cross SS, Brown NJ, Hamdy FC, Robson CN (2008) BMP-6 over-expression in prostate cancer is associated with increased Id-1 protein and a more invasive phenotype. J Pathol 214:394–404
David L, Mallet C, Mazerbourg S, Feige JJ, Bailly S (2007a) Identification of BMP9 and BMP10 as functional activators of the orphan activin receptor-like kinase 1 (ALK1) in endothelial cells. Blood 109:1953–1961
David L, Mallet C, Vailhe B, Lamouille S, Feige JJ, Bailly S (2007b) Activin receptor-like kinase 1 inhibits human microvascular endothelial cell migration: potential roles for JNK and ERK. J Cell Physiol 213:484–489
David L, Feige JJ, Bailly S (2009) Emerging role of bone morphogenetic proteins in angiogenesis. Cytokine Growth Factor Rev 20:203–212
de Jong DS, Steegenga WT, Hendriks JM, Zoelen EJ van, Olijve W, Dechering KJ (2004) Regulation of Notch signaling genes during BMP2-induced differentiation of osteoblast precursor cells. Biochem Biophys Res Commun 320:100–107
Deckers MM, Bezooijen RL van, Horst G van der, Hoogendam J, van Der Bent C, Papapoulos SE, Löwik CW (2002) Bone morphogenetic proteins stimulate angiogenesis through osteoblast-derived vascular endothelial growth factor A. Endocrinology 143:1545–1553
Dell’Accio F, De Bari C, El Tawil NM, Barone F, Mitsiadis TA, O’Dowd J, Pitzalis C (2006) Activation of WNT and BMP signaling in adult human articular cartilage following mechanical injury. Arthritis Res Ther 8:R139
Dorai H, Vukicevic S, Sampath TK (2000) Bone morphogenetic protein-7 (osteogenic protein-1) inhibits smooth muscle cell proliferation and stimulates the expression of markers that are characteristic of SMC phenotype in vitro. J Cell Physiol 184:37–45
D’Souza D, Patel K (1999) Involvement of long- and short-range signalling during early tendon development. Anat Embryol (Berl) 200:367–375
Ebrahem Q, Chaurasia SS, Vasanji A, Qi JH, Klenotic PA, Cutler A, Asosingh K, Erzurum S, Anand-Apte B (2010) Cross-talk between vascular endothelial growth factor and matrix metalloproteinases in the induction of neovascularization in vivo. Am J Pathol 176:496–503
Edom-Vovard F, Duprez D (2004) Signals regulating tendon formation during chick embryonic development. Dev Dyn 229:449–457
Edom-Vovard F, Bonnin M, Duprez D (2001) Fgf8 transcripts are located in tendons during embryonic chick limb development. Mech Dev 108:203–206
Einhorn TA, Majeska RJ, Mohaideen A, Kagel EM, Bouxsein ML, Turek TJ, Wozney JM (2003) A single percutaneous injection of recombinant human bone morphogenetic protein-2 accelerates fracture repair. J Bone Joint Surg Am 85-A:1425–1435
Ellis JE, Parker L, Cho J, Arora K (2010) Activin signaling functions upstream of Gbb to regulate synaptic growth at the Drosophila neuromuscular junction. Dev Biol 342:121–133
Endo M, Ohashi K, Mizuno K (2007) LIM kinase and slingshot are critical for neurite extension. J Biol Chem 282:13692–13702
Everman DB, Bartels CF, Yang Y, Yanamandra N, Goodman FR, Mendoza-Londono JR, Savarirayan R, White SM, Graham JM Jr, Gale RP, Svarch E, Newman WG, Kleckers AR, Francomano CA, Govindaiah V, Singh L, Morrison S, Thomas JT, Warman ML (2002) The mutational spectrum of brachydactyly type C. Am J Med Genet 112:291–296
Fang J, Zhu YY, Smiley E, Bonadio J, Rouleau JP, Goldstein SA, McCauley LK, Davidson BL, Roessler BJ (1996) Stimulation of new bone formation by direct transfer of osteogenic plasmid genes. Proc Natl Acad Sci USA 93:5753–5758
Fassbender M, Strobel C, Rauhe JS, Bergmann C, Schmidmaier G, Wildemann B (2011) Local inhibition of angiogenesis results in an atrophic non-union in a rat osteotomy model. Eur Cell Mater 22:1–11
Favier B, Rijli FM, Fromental-Ramain C, Fraulob V, Chambon P, Dolle P (1996) Functional cooperation between the non-paralogous genes Hoxa-10 and Hoxd-11 in the developing forelimb and axial skeleton. Development 122:449–460
Fazzalari NL (2011) Bone fracture and bone fracture repair. Osteoporos Int 22:2003–2006
Foletta VC, Lim MA, Soosairajah J, Kelly AP, Stanley EG, Shannon M, He W, Das S, Massague J, Bernard O (2003) Direct signaling by the BMP type II receptor via the cytoskeletal regulator LIMK1. J Cell Biol 162:1089–1098
Franceschi RT, Xiao G (2003) Regulation of the osteoblast-specific transcription factor, Runx2: responsiveness to multiple signal transduction pathways. J Cell Biochem 88:446–454
Francis-West PH, Abdelfattah A, Chen P, Allen C, Parish J, Ladher R, Allen S, MacPherson S, Luyten FP, Archer CW (1999) Mechanisms of GDF-5 action during skeletal development. Development 126:1305–1315
Franco M, Roswall P, Cortez E, Hanahan D, Pietras K (2011) Pericytes promote endothelial cell survival through induction of autocrine VEGF-A signaling and Bcl-w expression. Blood 118:2906–2917
Fu SC, Wong YP, Chan BP, Pau HM, Cheuk YC, Lee KM, Chan KM (2003) The roles of bone morphogenetic protein (BMP) 12 in stimulating the proliferation and matrix production of human patellar tendon fibroblasts. Life Sci 72:2965–2974
Fuentes-Medel Y, Budnik V (2010) Ménage à Trio during BMP-mediated retrograde signaling at the NMJ. Neuron 66:473–475
Gaengel K, Genove G, Armulik A, Betsholtz C (2009) Endothelial-mural cell signaling in vascular development and angiogenesis. Arterioscler Thromb Vasc Biol 29:630–638
Gamell C, Osses N, Bartrons R, Ruckle T, Camps M, Rosa JL, Ventura F (2008) BMP2 induction of actin cytoskeleton reorganization and cell migration requires PI3-kinase and Cdc42 activity. J Cell Sci 121:3960–3970
Gamell C, Susperregui AG, Bernard O, Rosa JL, Ventura F (2011) The p38/MK2/Hsp25 pathway is required for BMP-2-induced cell migration. PLoS One 6:e16477
Gerber C, Schneeberger AG, Perren SM, Nyffeler RW (1999) Experimental rotator cuff repair. A preliminary study. J Bone Joint Surg Am 81:1281–1290
Gerhardt H (2008) VEGF and endothelial guidance in angiogenic sprouting. Organogenesis 4:241–246
Geris L, Reed AA, Vander Sloten J, Simpson AH, Van Oosterwyck H (2010) Occurrence and treatment of bone atrophic non-unions investigated by an integrative approach. PLoS Comput Biol 6:e1000915
Ghosh-Choudhury N, Abboud SL, Nishimura R, Celeste A, Mahimainathan L, Choudhury GG (2002) Requirement of BMP-2-induced phosphatidylinositol 3-kinase and Akt serine/threonine kinase in osteoblast differentiation and Smad-dependent BMP-2 gene transcription. J Biol Chem 277:33361–33368
Glaser DL, Economides AN, Wang L, Liu X, Kimble RD, Fandl JP, Wilson JM, Stahl N, Kaplan FS, Shore EM (2003) In vivo somatic cell gene transfer of an engineered Noggin mutein prevents BMP4-induced heterotopic ossification. J Bone Joint Surg Am 85-A:2332–2342
Goold CP, Davis GW (2007) The BMP ligand Gbb gates the expression of synaptic homeostasis independent of synaptic growth control. Neuron 56:109–123
Graupera M, Guillermet-Guibert J, Foukas LC, Phng LK, Cain RJ, Salpekar A, Pearce W, Meek S, Millan J, Cutillas PR, Smith AJ, Ridley AJ, Ruhrberg C, Gerhardt H, Vanhaesebroeck B (2008) Angiogenesis selectively requires the p110alpha isoform of PI3K to control endothelial cell migration. Nature 453:662–666
Greenblatt MB, Shim JH, Zou W, Sitara D, Schweitzer M, Hu D, Lotinun S, Sano Y, Baron R, Park JM, Arthur S, Xie M, Schneider MD, Zhai B, Gygi S, Davis R, Glimcher LH (2010) The p38 MAPK pathway is essential for skeletogenesis and bone homeostasis in mice. J Clin Invest 120:2457–2473
Gruber R, Karreth F, Fischer MB, Watzek G (2002) Platelet-released supernatants stimulate formation of osteoclast-like cells through a prostaglandin/RANKL-dependent mechanism. Bone 30:726–732
Gruneberg H, Lee AJ (1973) The anatomy and development of brachypodism in the mouse. J Embryol Exp Morphol 30:119–141
Guicheux J, Lemonnier J, Ghayor C, Suzuki A, Palmer G, Caverzasio J (2003) Activation of p38 mitogen-activated protein kinase and c-Jun-NH2-terminal kinase by BMP-2 and their implication in the stimulation of osteoblastic cell differentiation. J Bone Miner Res 18:2060–2068
Halbrooks PJ, Ding R, Wozney JM, Bain G (2007) Role of RGM coreceptors in bone morphogenetic protein signaling. J Mol Signal 2:4
Hall A (1998) Rho GTPases and the actin cytoskeleton. Science 279:509–514
Hall AK, Miller RH (2004) Emerging roles for bone morphogenetic proteins in central nervous system glial biology. J Neurosci Res 76:1–8
Hankenson KD, Dishowitz M, Gray C, Schenker M (2011) Angiogenesis in bone regeneration. Injury 42:556–561
Hannallah D, Peng H, Young B, Usas A, Gearhart B, Huard J (2004) Retroviral delivery of Noggin inhibits the formation of heterotopic ossification induced by BMP-4, demineralized bone matrix, and trauma in an animal model. J Bone Joint Surg Am 86-A:80–91
Hansmann G, Jesus Perez V de, Alastalo TP, Alvira CM, Guignabert C, Bekker JM, Schellong S, Urashima T, Wang L, Morrell NW, Rabinovitch M (2008) An antiproliferative BMP-2/PPARgamma/apoE axis in human and murine SMCs and its role in pulmonary hypertension. J Clin Invest 118:1846–1857
Hao J, Ho JN, Lewis JA, Karim KA, Daniels RN, Gentry PR, Hopkins CR, Lindsley CW, Hong CC (2010) In vivo structure-activity relationship study of dorsomorphin analogues identifies selective VEGF and BMP inhibitors. ACS Chem Biol 5:245–253
Heinke J, Wehofsits L, Zhou Q, Zoeller C, Baar KM, Helbing T, Laib A, Augustin H, Bode C, Patterson C, Moser M (2008) BMPER is an endothelial cell regulator and controls bone morphogenetic protein-4-dependent angiogenesis. Circ Res 103:804–812
Hellberg C, Ostman A, Heldin CH (2010) PDGF and vessel maturation. Recent Results Cancer Res 180:103–114
Hellstrom M, Phng LK, Hofmann JJ, Wallgard E, Coultas L, Lindblom P, Alva J, Nilsson AK, Karlsson L, Gaiano N, Yoon K, Rossant J, Iruela-Arispe ML, Kalen M, Gerhardt H, Betsholtz C (2007) Dll4 signalling through Notch1 regulates formation of tip cells during angiogenesis. Nature 445:776–780
Helm GA, Alden TD, Sheehan JP, Kallmes D (2000) Bone morphogenetic proteins and bone morphogenetic protein gene therapy in neurological surgery: a review. Neurosurgery 46:1213–1222
Henriksen K, Karsdal M, Delaisse JM, Engsig MT (2003) RANKL and vascular endothelial growth factor (VEGF) induce osteoclast chemotaxis through an ERK1/2-dependent mechanism. J Biol Chem 278:48745–48753
Higashikawa A, Kawaguchi H, Nakamura K, Chung U (2006) Interactions of chondrocytes and osteoblasts during endochondral bone formation. Clin Calcium 16:829–836
Hilton MJ, Tu X, Wu X, Bai S, Zhao H, Kobayashi T, Kronenberg HM, Teitelbaum SL, Ross FP, Kopan R, Long F (2008) Notch signaling maintains bone marrow mesenchymal progenitors by suppressing osteoblast differentiation. Nat Med 14:306–314
Hoffmann A, Pelled G, Turgeman G, Eberle P, Zilberman Y, Shinar H, Keinan-Adamsky K, Winkel A, Shahab S, Navon G, Gross G, Gazit D (2006) Neotendon formation induced by manipulation of the Smad8 signalling pathway in mesenchymal stem cells. J Clin Invest 116:940–952
Holderfield MT, Hughes CC (2008) Crosstalk between vascular endothelial growth factor, notch, and transforming growth factor-beta in vascular morphogenesis. Circ Res 102:637–652
Holley SA, Jackson PD, Sasai Y, Lu B, De Robertis EM, Hoffmann FM, Ferguson EL (1995) A conserved system for dorsal-ventral patterning in insects and vertebrates involving sog and chordin. Nature 376:249–253
Horwitz EM (2010) Building bone from blood vessels. Nat Med 16:1373–1374
Husmann I, Soulet L, Gautron J, Martelly I, Barritault D (1996) Growth factors in skeletal muscle regeneration. Cytokine Growth Factor Rev 7:249–258
Itoh F, Itoh S, Goumans MJ, Valdimarsdottir G, Iso T, Dotto GP, Hamamori Y, Kedes L, Kato M, Dijke P ten (2004) Synergy and antagonism between Notch and BMP receptor signaling pathways in endothelial cells. EMBO J 23:541–551
Jozsa L, Reffy A, Kannus P, Demel S, Elek E (1990) Pathological alterations in human tendons. Arch Orthop Trauma Surg 110:15–21
Kang Q, Sun MH, Cheng H, Peng Y, Montag AG, Deyrup AT, Jiang W, Luu HH, Luo J, Szatkowski JP, Vanichakarn P, Park JY, Li Y, Haydon RC, He TC (2004) Characterization of the distinct orthotopic bone-forming activity of 14 BMPs using recombinant adenovirus-mediated gene delivery. Gene Ther 11:1312–1320
Kannus P (2000) Structure of the tendon connective tissue. Scand J Med Sci Sports 10:312–320
Kasper G, Glaeser JD, Geissler S, Ode A, Tuischer J, Matziolis G, Perka C, Duda GN (2007) Matrix metalloprotease activity is an essential link between mechanical stimulus and mesenchymal stem cell behavior. Stem Cells 25:1985–1994
Kelley R, Ren R, Pi X, Wu Y, Moreno I, Willis M, Moser M, Ross M, Podkowa M, Attisano L, Patterson C (2009) A concentration-dependent endocytic trap and sink mechanism converts Bmper from an activator to an inhibitor of Bmp signaling. J Cell Biol 184:597–609
Kempen DH, Lu L, Heijink A, Hefferan TE, Creemers LB, Maran A, Yaszemski MJ, Dhert WJ (2009) Effect of local sequential VEGF and BMP-2 delivery on ectopic and orthotopic bone regeneration. Biomaterials 30:2816–2825
Kim YJ, Lee MH, Wozney JM, Cho JY, Ryoo HM (2004) Bone morphogenetic protein-2-induced alkaline phosphatase expression is stimulated by Dlx5 and repressed by Msx2. J Biol Chem 279:50773–50780
Kirkpatrick CJ, Fuchs S, Unger RE (2011) Co-culture systems for vascularization—learning from nature. Adv Drug Deliv Rev 63:291–299
Kodaira K, Imada M, Goto M, Tomoyasu A, Fukuda T, Kamijo R, Suda T, Higashio K, Katagiri T (2006) Purification and identification of a BMP-like factor from bovine serum. Biochem Biophys Res Commun 345:1224–1231
Kohno T, Ishibashi Y, Tsuda E, Kusumi T, Tanaka M, Toh S (2007) Immunohistochemical demonstration of growth factors at the tendon-bone interface in anterior cruciate ligament reconstruction using a rabbit model. J Orthop Sci 12:67–73
Kolar P, Schmidt-Bleek K, Schell H, Gaber T, Toben D, Schmidmaier G, Perka C, Buttgereit F, Duda GN (2010) The early fracture hematoma and its potential role in fracture healing. Tissue Eng Part B Rev 16:427–434
Kolar P, Gaber T, Perka C, Duda GN, Buttgereit F (2011) Human early fracture hematoma is characterized by inflammation and hypoxia. Clin Orthop Relat Res 469:3118-3126
Koleva RI, Conley BA, Romero D, Riley KS, Marto JA, Lux A, Vary CP (2006) Endoglin structure and function: determinants of endoglin phosphorylation by transforming growth factor-beta receptors. J Biol Chem 281:25110–25123
Krause C, Guzman A, Knaus P (2011) Noggin. Int J Biochem Cell Biol 43:478–481
Kuang S, Kuroda K, Le GF, Rudnicki MA (2007) Asymmetric self-renewal and commitment of satellite stem cells in muscle. Cell 129:999–1010
Kuang S, Gillespie MA, Rudnicki MA (2008) Niche regulation of muscle satellite cell self-renewal and differentiation. Cell Stem Cell 2:22–31
Kumar D, Shadrach JL, Wagers AJ, Lassar AB (2009) Id3 is a direct transcriptional target of Pax7 in quiescent satellite cells. Mol Biol Cell 20:3170–3177
Kuroda R, Usas A, Kubo S, Corsi K, Peng H, Rose T, Cummins J, Fu FH, Huard J (2006) Cartilage repair using bone morphogenetic protein 4 and muscle-derived stem cells. Arthritis Rheum 54:433–442
Kuroda R, Ishida K, Matsumoto T, Akisue T, Fujioka H, Mizuno K, Ohgushi H, Wakitani S, Kurosaka M (2007) Treatment of a full-thickness articular cartilage defect in the femoral condyle of an athlete with autologous bone-marrow stromal cells. Osteoarthritis Cartilage 15:226–231
Lechleider RJ, Ryan JL, Garrett L, Eng C, Deng C, Wynshaw-Boris A, Roberts AB (2001) Targeted mutagenesis of Smad1 reveals an essential role in chorioallantoic fusion. Dev Biol 240:157–167
Lee JY, Zhou Z, Taub PJ, Ramcharan M, Li Y, Akinbiyi T, Maharam ER, Leong DJ, Laudier DM, Ruike T, Torina PJ, Zaidi M, Majeska RJ, Schaffler MB, Flatow EL, Sun HB (2011) BMP-12 treatment of adult mesenchymal stem cells in vitro augments tendon-like tissue formation and defect repair in vivo. PLoS One 6:e17531
Lehmann K, Seemann P, Stricker S, Sammar M, Meyer B, Suring K, Majewski F, Tinschert S, Grzeschik KH, Muller D, Knaus P, Nurnberg P, Mundlos S (2003) Mutations in bone morphogenetic protein receptor 1B cause brachydactyly type A2. Proc Natl Acad Sci USA 100:12277–12282
Lehmann W, Edgar CM, Wang K, Cho TJ, Barnes GL, Kakar S, Graves DT, Rueger JM, Gerstenfeld LC, Einhorn TA (2005) Tumor necrosis factor alpha (TNF-alpha) coordinately regulates the expression of specific matrix metalloproteinases (MMPS) and angiogenic factors during fracture healing. Bone 36:300–310
Li DY, Sorensen LK, Brooke BS, Urness LD, Davis EC, Taylor DG, Boak BB, Wendel DP (1999) Defective angiogenesis in mice lacking endoglin. Science 284:1534–1537
Lienau J, Schmidt-Bleek K, Peters A, Haschke F, Duda GN, Perka C, Bail HJ, Schutze N, Jakob F, Schell H (2009) Differential regulation of blood vessel formation between standard and delayed bone healing. J Orthop Res 27:1133–1140
Lieu S, Hansen E, Dedini R, Behonick D, Werb Z, Miclau T, Marcucio R, Colnot C (2011) Impaired remodeling phase of fracture repair in the absence of matrix metalloproteinase-2. Dis Model Mech 4:203–211
Liu A, Niswander LA (2005) Bone morphogenetic protein signalling and vertebrate nervous system development. Nat Rev Neurosci 6:945–954
Liu J, Wilson S, Reh T (2003) BMP receptor 1b is required for axon guidance and cell survival in the developing retina. Dev Biol 256:34–48
Liu Z, Shen J, Pu K, Katus HA, Ploger F, Tiefenbacher CP, Chen X, Braun T (2009) GDF5 and BMP2 inhibit apoptosis via activation of BMPR2 and subsequent stabilization of XIAP. Biochim Biophys Acta 1793:1819–1827
Lou J, Tu Y, Burns M, Silva MJ, Manske P (2001) BMP-12 gene transfer augmentation of lacerated tendon repair. J Orthop Res 19:1199–1202
Lu C, Miclau T, Hu D, Marcucio RS (2007) Ischemia leads to delayed union during fracture healing: a mouse model. J Orthop Res 25:51–61
Luu HH, Song WX, Luo X, Manning D, Luo J, Deng ZL, Sharff KA, Montag AG, Haydon RC, He TC (2007) Distinct roles of bone morphogenetic proteins in osteogenic differentiation of mesenchymal stem cells. J Orthop Res 25:665–677
Maeda Y, Tsuji K, Nifuji A, Noda M (2004) Inhibitory helix-loop-helix transcription factors Id1/Id3 promote bone formation in vivo. J Cell Biochem 93:337–344
Maes C, Stockmans I, Moermans K, Van LR, Smets N, Carmeliet P, Bouillon R, Carmeliet G (2004) Soluble VEGF isoforms are essential for establishing epiphyseal vascularization and regulating chondrocyte development and survival. J Clin Invest 113:188–199
Maes C, Kobayashi T, Selig MK, Torrekens S, Roth SI, Mackem S, Carmeliet G, Kronenberg HM (2010) Osteoblast precursors, but not mature osteoblasts, move into developing and fractured bones along with invading blood vessels. Dev Cell 19:329–344
Maffulli N, Wong J, Almekinders LC (2003) Types and epidemiology of tendinopathy. Clin Sports Med 22:675–692
Martinek V, Latterman C, Usas A, Abramowitch S, Woo SL, Fu FH, Huard J (2002) Enhancement of tendon-bone integration of anterior cruciate ligament grafts with bone morphogenetic protein-2 gene transfer: a histological and biomechanical study. J Bone Joint Surg Am 84-A:1123–1131
Massague J, Seoane J, Wotton D (2005) Smad transcription factors. Genes Dev 19:2783–2810
Matsuguchi T, Chiba N, Bandow K, Kakimoto K, Masuda A, Ohnishi T (2009) JNK activity is essential for Atf4 expression and late-stage osteoblast differentiation. J Bone Miner Res 24:398–410
Matsumoto T, Mifune Y, Kawamoto A, Kuroda R, Shoji T, Iwasaki H, Suzuki T, Oyamada A, Horii M, Yokoyama A, Nishimura H, Lee SY, Miwa M, Doita M, Kurosaka M, Asahara T (2008) Fracture induced mobilization and incorporation of bone marrow-derived endothelial progenitor cells for bone healing. J Cell Physiol 215:234–242
Matsuura I, Taniguchi J, Hata K, Saeki N, Yamashita T (2008) BMP inhibition enhances axonal growth and functional recovery after spinal cord injury. J Neurochem 105:1471–1479
Matuszewski PE, Mehta S (2011) Fracture consolidation in a tibial nonunion after revascularization: a case report. J Orthop Trauma 25:e15–e20
McCabe BD, Marques G, Haghighi AP, Fetter RD, Crotty ML, Haerry TE, Goodman CS, O’Connor MB (2003) The BMP homolog Gbb provides a retrograde signal that regulates synaptic growth at the Drosophila neuromuscular junction. Neuron 39:241–254
Meberg PJ (2000) Signal-regulated ADF/cofilin activity and growth cone motility. Mol Neurobiol 21:97–107
Medici D, Shore EM, Lounev VY, Kaplan FS, Kalluri R, Olsen BR (2010) Conversion of vascular endothelial cells into multipotent stem-like cells. Nat Med 16:1400–1406
Medley TL, Cole TJ, Dart AM, Gatzka CD, Kingwell BA (2004) Matrix metalloproteinase-9 genotype influences large artery stiffness through effects on aortic gene and protein expression. Arterioscler Thromb Vasc Biol 24:1479–1484
Mikic B (2004) Multiple effects of GDF-5 deficiency on skeletal tissues: implications for therapeutic bioengineering. Ann Biomed Eng 32:466–476
Mikic B, Rossmeier K, Bierwert L (2009a) Identification of a tendon phenotype in GDF6 deficient mice. Anat Rec 292:396–400
Mikic B, Rossmeier K, Bierwert L (2009b) Sexual dimorphism in the effect of GDF-6 deficiency on murine tendon. J Orthop Res 27:1603–1611
Mitola S, Ravelli C, Moroni E, Salvi V, Leali D, Ballmer-Hofer K, Zammataro L, Presta M (2010) Gremlin is a novel agonist of the major proangiogenic receptor VEGFR2. Blood 116:3677–3680
Miyazawa K, Shinozaki M, Hara T, Furuya T, Miyazono K (2002) Two major Smad pathways in TGF-beta superfamily signalling. Genes Cells 7:1191–1204
Miyazono K, Maeda S, Imamura T (2005) BMP receptor signaling: transcriptional targets, regulation of signals, and signaling cross-talk. Cytokine Growth Factor Rev 16:251–263
Morikawa M, Koinuma D, Tsutsumi S, Vasilaki E, Kanki Y, Heldin CH, Aburatani H, Miyazono K (2011) ChIP-seq reveals cell type-specific binding patterns of BMP-specific Smads and a novel binding motif. Nucleic Acids Res 39:8712–8727
Morrell NW, Yang X, Upton PD, Jourdan KB, Morgan N, Sheares KK, Trembath RC (2001) Altered growth responses of pulmonary artery smooth muscle cells from patients with primary pulmonary hypertension to transforming growth factor-beta(1) and bone morphogenetic proteins. Circulation 104:790–795
Moser M, Patterson C (2005) Bone morphogenetic proteins and vascular differentiation: BMPing up vasculogenesis. Thromb Haemost 94:713–718
Moser M, Binder O, Wu Y, Aitsebaomo J, Ren R, Bode C, Bautch VL, Conlon FL, Patterson C (2003) BMPER, a novel endothelial cell precursor-derived protein, antagonizes bone morphogenetic protein signaling and endothelial cell differentiation. Mol Cell Biol 23:5664–5679
Mounier R, Chretien F, Chazaud B (2011) Blood vessels and the satellite cell niche. Curr Top Dev Biol 96:121–138
Mu Y, Gudey SK, Landstrom M (2011) Non-Smad signaling pathways. Cell Tissue Res. doi:10.1007/s00441-011-1201-y
Mukherjee A, Rotwein P (2009) Akt promotes BMP2-mediated osteoblast differentiation and bone development. J Cell Sci 122:716–726
Nakagawa Y, Ikeda K, Akakabe Y, Koide M, Uraoka M, Yutaka KT, Kurimoto-Nakano R, Takahashi T, Matoba S, Yamada H, Okigaki M, Matsubara H (2010) Paracrine osteogenic signals via bone morphogenetic protein-2 accelerate the atherosclerotic intimal calcification in vivo. Arterioscler Thromb Vasc Biol 30:1908–1915
Nakashima K, Takizawa T, Ochiai W, Yanagisawa M, Hisatsune T, Nakafuku M, Miyazono K, Kishimoto T, Kageyama R, Taga T (2001) BMP2-mediated alteration in the developmental pathway of fetal mouse brain cells from neurogenesis to astrocytogenesis. Proc Natl Acad Sci USA 98:5868–5873
Nauth A, Miclau T III, Li R, Schemitsch EH (2010) Gene therapy for fracture healing. J Orthop Trauma 24 (Suppl 1):S17–S24
Neuhaus H, Rosen V, Thies RS (1999) Heart specific expression of mouse BMP-10 a novel member of the TGF-beta superfamily. Mech Dev 80:181–184
Niikura T, Hak DJ, Reddi AH (2006) Global gene profiling reveals a downregulation of BMP gene expression in experimental atrophic nonunions compared to standard healing fractures. J Orthop Res 24:1463–1471
Nilsson I, Bahram F, Li X, Gualandi L, Koch S, Jarvius M, Soderberg O, Anisimov A, Kholova I, Pytowski B, Baldwin M, Yla-Herttuala S, Alitalo K, Kreuger J, Claesson-Welsh L (2010) VEGF receptor 2/-3 heterodimers detected in situ by proximity ligation on angiogenic sprouts. EMBO J 29:1377–1388
Nishimura R, Hata K, Harris SE, Ikeda F, Yoneda T (2002) Core-binding factor alpha 1 (Cbfa1) induces osteoblastic differentiation of C2C12 cells without interactions with Smad1 and Smad5. Bone 31:303–312
Nishitoh H, Ichijo H, Kimura M, Matsumoto T, Makishima F, Yamaguchi A, Yamashita H, Enomoto S, Miyazono K (1996) Identification of type I and type II serine/threonine kinase receptors for growth/differentiation factor-5. J Biol Chem 271:21345–21352
Nojima J, Kanomata K, Takada Y, Fukuda T, Kokabu S, Ohte S, Takada T, Tsukui T, Yamamoto TS, Sasanuma H, Yoneyama K, Ueno N, Okazaki Y, Kamijo R, Yoda T, Katagiri T (2010) Dual roles of smad proteins in the conversion from myoblasts to osteoblastic cells by bone morphogenetic proteins. J Biol Chem 285:15577–15586
O’Connor-Giles KM, Ho LL, Ganetzky B (2008) Nervous wreck interacts with thickveins and the endocytic machinery to attenuate retrograde BMP signaling during synaptic growth. Neuron 58:507–518
O’Driscoll SW (1998) The healing and regeneration of articular cartilage. J Bone Joint Surg Am 80:1795–1812
Oh SP, Seki T, Goss KA, Imamura T, Yi Y, Donahoe PK, Li L, Miyazono K, Dijke P ten, Kim S, Li E (2000) Activin receptor-like kinase 1 modulates transforming growth factor-beta 1 signaling in the regulation of angiogenesis. Proc Natl Acad Sci USA 97:2626–2631
Ohashi K, Hosoya T, Takahashi K, Hing H, Mizuno K (2000) A Drosophila homolog of LIM-kinase phosphorylates cofilin and induces actin cytoskeletal reorganization. Biochem Biophys Res Commun 276:1178–1185
Ono Y, Calhabeu F, Morgan JE, Katagiri T, Amthor H, Zammit PS (2011) BMP signalling permits population expansion by preventing premature myogenic differentiation in muscle satellite cells. Cell Death Differ 18:222–234
Ortuno MJ, Ruiz-Gaspa S, Rodriguez-Carballo E, Susperregui AR, Bartrons R, Rosa JL, Ventura F (2010) p38 regulates expression of osteoblast-specific genes by phosphorylation of osterix. J Biol Chem 285:31985–31994
Oshin AO, Stewart MC (2007) The role of bone morphogenetic proteins in articular cartilage development, homeostasis and repair. Vet Comp Orthop Traumatol 20:151–158
Park EJ, Kim ES, Weber HP, Wright RF, Mooney DJ (2008) Improved bone healing by angiogenic factor-enriched platelet-rich plasma and its synergistic enhancement by bone morphogenetic protein-2. Int J Oral Maxillofac Implants 23:818–826
Patil AS, Sable RB, Kothari RM (2011) Occurrence, biochemical profile of vascular endothelial growth factor (VEGF) isoforms and their functions in endochondral ossification. J Cell Physiol. doi:10.1002/jcp.22846
Pauly S, Klatte F, Strobel C, Schmidmaier G, Greiner S, Scheibel M, Wildemann B (2011) BMP-2 and BMP-7 affect human rotator cuff tendon cells in vitro. J Shoulder Elbow Surg (in press)
Payne KA, Meszaros LB, Phillippi JA, Huard J (2010) Effect of phosphatidyl inositol 3-kinase, extracellular signal-regulated kinases 1/2, and p38 mitogen-activated protein kinase inhibition on osteogenic differentiation of muscle-derived stem cells. Tissue Eng Part A 16:3647–3655
Peng XD, Xu PZ, Chen ML, Hahn-Windgassen A, Skeen J, Jacobs J, Sundararajan D, Chen WS, Crawford SE, Coleman KG, Hay N (2003) Dwarfism, impaired skin development, skeletal muscle atrophy, delayed bone development, and impeded adipogenesis in mice lacking Akt1 and Akt2. Genes Dev 17:1352–1365
Peng H, Usas A, Olshanski A, Ho AM, Gearhart B, Cooper GM, Huard J (2005) VEGF improves, whereas sFlt1 inhibits, BMP2-induced bone formation and bone healing through modulation of angiogenesis. J Bone Miner Res 20:2017–2027
Polinkovsky A, Robin NH, Thomas JT, Irons M, Lynn A, Goodman FR, Reardon W, Kant SG, Brunner HG, Burgt I van der, Chitayat D, McGaughran J, Donnai D, Luyten FP, Warman ML (1997) Mutations in CDMP1 cause autosomal dominant brachydactyly type C. Nat Genet 17:18–19
Proschel C, Blouin MJ, Gutowski NJ, Ludwig R, Noble M (1995) Limk1 is predominantly expressed in neural tissues and phosphorylates serine, threonine and tyrosine residues in vitro. Oncogene 11:1271–1281
Raida M, Heymann AC, Gunther C, Niederwieser D (2006) Role of bone morphogenetic protein 2 in the crosstalk between endothelial progenitor cells and mesenchymal stem cells. Int J Mol Med 18:735–739
Raucci A, Bellosta P, Grassi R, Basilico C, Mansukhani A (2008) Osteoblast proliferation or differentiation is regulated by relative strengths of opposing signaling pathways. J Cell Physiol 215:442–451
Reddi AH (2005) BMPs: from bone morphogenetic proteins to body morphogenetic proteins. Cytokine Growth Factor Rev 16:249–250
Reshef R, Maroto M, Lassar AB (1998) Regulation of dorsal somitic cell fates: BMPs and Noggin control the timing and pattern of myogenic regulator expression. Genes Dev 12:290–303
Reumann MK, Nair T, Strachna O, Boskey AL, Mayer-Kuckuk P (2010) Production of VEGF receptor 1 and 2 mRNA and protein during endochondral bone repair is differential and healing phase specific. J Appl Physiol 109:1930–1938
Ribatti D, Nico B, Crivellato E (2011) The role of pericytes in angiogenesis. Int J Dev Biol 55:261–268
Rifas L (2006) T-cell cytokine induction of BMP-2 regulates human mesenchymal stromal cell differentiation and mineralization. J Cell Biochem 98:706–714
Rodeo SA, Arnoczky SP, Torzilli PA, Hidaka C, Warren RF (1993) Tendon-healing in a bone tunnel. A biomechanical and histological study in the dog. J Bone Joint Surg Am 75:1795–1803
Ronzière MC, Aubert-Foucher E, Gouttenoire J, Bernaud J, Herbage D, Mallein-Gerin F (2005) Integrin alpha1beta1 mediates collagen induction of MMP-13 expression in MC615 chondrocytes. Biochim Biophys Acta 1746:55–64
Ros MA, Rivero FB, Hinchliffe JR, Hurle JM (1995) Immunohistological and ultrastructural study of the developing tendons of the avian foot. Anat Embryol (Berl) 192:483–496
Rountree RB, Schoor M, Chen H, Marks ME, Harley V, Mishina Y, Kingsley DM (2004) BMP receptor signaling is required for postnatal maintenance of articular cartilage. PLoS Biol 2:e355
Rudnicki MA, Le Grand F, McKinnell I, Kuang S (2008) The molecular regulation of muscle stem cell function. Cold Spring Harb Symp Quant Biol 73:323–331
Sabo JK, Kilpatrick TJ, Cate HS (2009) Effects of bone morphogenic proteins on neural precursor cells and regulation during central nervous system injury. Neurosignals 17:255–264
Salinas S, Bilsland LG, Schiavo G (2008) Molecular landmarks along the axonal route: axonal transport in health and disease. Curr Opin Cell Biol 20:445–453
Sammar M, Stricker S, Schwabe GC, Sieber C, Hartung A, Hanke M, Oishi I, Pohl J, Minami Y, Sebald W, Mundlos S, Knaus P (2004) Modulation of GDF5/BRI-b signalling through interaction with the tyrosine kinase receptor Ror2. Genes Cells 9:1227–1238
Scharpfenecker M, Dinther M van, Liu Z, Bezooijen RL van, Zhao Q, Pukac L, Löwik CW, Dijke P ten (2007) BMP-9 signals via ALK1 and inhibits bFGF-induced endothelial cell proliferation and VEGF-stimulated angiogenesis. J Cell Sci 120:964–972
Schindowski K, Bohlen und Halback O von, Strelau J, Ridder DA, Herrmann O, Schober A, Schwaninger M, Unsicker K (2011) Regulation of GDF-15, a distant TGF-beta superfamily member, in a mouse model of cerebral ischemia. Cell Tissue Res 343:399–409
Schluesener HJ, Meyermann R (1995) Immunolocalization of BMP-6, a novel TGF-beta-related cytokine, in normal and atherosclerotic smooth muscle cells. Atherosclerosis 113:153–156
Schmidt-Bleek K, Schell H, Schulz N, Hoff P, Perka C, Buttgereit F, Volk HD, Lienau J, Duda GN (2011) Inflammatory phase of bone healing initiates the regenerative healing cascade. Cell Tissue Res (in press)
Schwaerzer GK, Hiepen C, Schrewe H, Nickel J, Ploeger F, Sebald W, Mueller T, Knaus P (2011) New insights into the molecular mechanism of multiple synostoses syndrome (SYNS): mutation within the GDF5 knuckle epitope causes noggin-resistance. J Bone Miner Res. doi:10.1002/jbmr.532
Schweitzer R, Chyung JH, Murtaugh LC, Brent AE, Rosen V, Olson EN, Lassar A, Tabin CJ (2001) Analysis of the tendon cell fate using Scleraxis, a specific marker for tendons and ligaments. Development 128:3855–3866
Seale P, Sabourin LA, Girgis-Gabardo A, Mansouri A, Gruss P, Rudnicki MA (2000) Pax7 is required for the specification of myogenic satellite cells. Cell 102:777–786
See J, Mamontov P, Ahn K, Wine-Lee L, Crenshaw EB III, Grinspan JB (2007) BMP signaling mutant mice exhibit glial cell maturation defects. Mol Cell Neurosci 35:171–182
Seemann P, Brehm A, König J, Reissner C, Stricker S, Kuss P, Haupt J, Renninger S, Nickel J, Sebald W, Groppe JC, Plöger F, Pohl J, Schmidt-von Kegler M, Walther M, Gassner I, Rusu C, Janecke AR, Dathe K, Mundlos S (2009) Mutations in GDF5 reveal a key residue mediating BMP inhibition by NOGGIN. PLoS Genet 5:e1000747
Seib FP, Lanfer B, Bornhauser M, Werner C (2010) Biological activity of extracellular matrix-associated BMP-2. J Tissue Eng Regen Med 4:324–327
Setoguchi T, Yone K, Matsuoka E, Takenouchi H, Nakashima K, Sakou T, Komiya S, Izumo S (2001) Traumatic injury-induced BMP7 expression in the adult rat spinal cord. Brain Res 921:219–225
Shah AK, Lazatin J, Sinha RK, Lennox T, Hickok NJ, Tuan RS (1999) Mechanism of BMP-2 stimulated adhesion of osteoblastic cells to titanium alloy. Biol Cell 91:131–142
Sieber C, Kopf J, Hiepen C, Knaus P (2009) Recent advances in BMP receptor signaling. Cytokine Growth Factor Rev 20:343–355
Song X, Liu S, Qu X, Hu Y, Zhang X, Wang T, Wei F (2011) BMP2 and VEGF promote angiogenesis but retard terminal differentiation of osteoblasts in bone regeneration by up-regulating Id1. Acta Biochim Biophys Sin (Shanghai) 43:796–804
Soosairajah J, Maiti S, Wiggan O, Sarmiere P, Moussi N, Sarcevic B, Sampath R, Bamburg JR, Bernard O (2005) Interplay between components of a novel LIM kinase-slingshot phosphatase complex regulates cofilin. EMBO J 24:473–486
Sorrentino A, Thakur N, Grimsby S, Marcusson A, Bulow V von, Schuster N, Zhang S, Heldin CH, Landstrom M (2008) The type I TGF-beta receptor engages TRAF6 to activate TAK1 in a receptor kinase-independent manner. Nat Cell Biol 10:1199–1207
Stabile H, Mitola S, Moroni E, Belleri M, Nicoli S, Coltrini D, Peri F, Pessi A, Orsatti L, Talamo F, Castronovo V, Waltregny D, Cotelli F, Ribatti D, Presta M (2007) Bone morphogenic protein antagonist Drm/gremlin is a novel proangiogenic factor. Blood 109:1834–1840
Steinert A, Weber M, Dimmler A, Julius C, Schutze N, Noth U, Cramer H, Eulert J, Zimmermann U, Hendrich C (2003) Chondrogenic differentiation of mesenchymal progenitor cells encapsulated in ultrahigh-viscosity alginate. J Orthop Res 21:1090–1097
Steinwachs MR, Guggi T, Kreuz PC (2008) Marrow stimulation techniques. Injury 39 (Suppl 1):S26–S31
Sterrenburg E, Wees CG van der, White SJ, Turk R, Menezes RX de, Ommen GJ van, Dunnen JT den, t’Hoen PA (2006) Gene expression profiling highlights defective myogenesis in DMD patients and a possible role for bone morphogenetic protein 4. Neurobiol Dis 23:228–236
Storm EE, Huynh TV, Copeland NG, Jenkins NA, Kingsley DM, Lee SJ (1994) Limb alterations in brachypodism mice due to mutations in a new member of the TGF beta-superfamily. Nature 368:639–643
Strelau J, Sullivan A, Bottner M, Lingor P, Falkenstein E, Suter-Crazzolara C, Galter D, Jaszai J, Krieglstein K, Unsicker K (2000) Growth/differentiation factor-15/macrophage inhibitory cytokine-1 is a novel trophic factor for midbrain dopaminergic neurons in vivo. J Neurosci 20:8597–8603
Strelau J, Schober A, Sullivan A, Schilling L, Unsicker K (2003) Growth/differentiation factor-15 (GDF-15), a novel member of the TGF-beta superfamily, promotes survival of lesioned mesencephalic dopaminergic neurons in vitro and in vivo and is induced in neurons following cortical lesioning. J Neural Transm Suppl 65:197-203
Strelau J, Strzelczyk A, Rusu P, Bendner G, Wiese S, Diella F, Altick AL, Bartheld CS von, Klein R, Sendtner M, Unsicker K (2009) Progressive postnatal motoneuron loss in mice lacking GDF-15. J Neurosci 29:13640–13648
Su Y, Zheng L, Wang Q, Bao J, Cai Z, Liu A (2010) The PI3K/Akt pathway upregulates Id1 and integrin alpha4 to enhance recruitment of human ovarian cancer endothelial progenitor cells. BMC Cancer 10:459
Subramaniam S, Strelau J, Unsicker K (2003) Growth differentiation factor-15 prevents low potassium-induced cell death of cerebellar granule neurons by differential regulation of Akt and ERK pathways. J Biol Chem 278:8904–8912
Suchting S, Freitas C, Noble F le, Benedito R, Bréant C, Duarte A, Eichmann A (2007) The Notch ligand Delta-like 4 negatively regulates endothelial tip cell formation and vessel branching. Proc Natl Acad Sci USA 104:3225–3230
Suzuki Y, Montagne K, Nishihara A, Watabe T, Miyazono K (2008) BMPs promote proliferation and migration of endothelial cells via stimulation of VEGF-A/VEGFR2 and angiopoietin-1/Tie2 signalling. J Biochem 143:199–206
Suzuki Y, Ohga N, Morishita Y, Hida K, Miyazono K, Watabe T (2010) BMP-9 induces proliferation of multiple types of endothelial cells in vitro and in vivo. J Cell Sci 123:1684–1692
Takizawa T, Ochiai W, Nakashima K, Taga T (2003) Enhanced gene activation by Notch and BMP signaling cross-talk. Nucleic Acids Res 31:5723–5731
Taylor KL, Henderson AM, Hughes CC (2002) Notch activation during endothelial cell network formation in vitro targets the basic HLH transcription factor HESR-1 and downregulates VEGFR-2/KDR expression. Microvasc Res 64:372–383
Tian F, Zhou AX, Smits AM, Larsson E, Goumans MJ, Heldin CH, Boren J, Akyurek LM (2010) Endothelial cells are activated during hypoxia via endoglin/ALK-1/SMAD1/5 signaling in vivo and in vitro. Biochem Biophys Res Commun 392:283–288
Towler DA, Gelberman RH (2006) The alchemy of tendon repair: a primer for the (S)mad scientist. J Clin Invest 116:863–866
Tseng WP, Yang SN, Lai CH, Tang CH (2010) Hypoxia induces BMP-2 expression via ILK, Akt, mTOR, and HIF-1 pathways in osteoblasts. J Cell Physiol 223:810–818
Tsubone T, Moran SL, Amadio PC, Zhao C, An KN (2004) Expression of growth factors in canine flexor tendon after laceration in vivo. Ann Plast Surg 53:393–397
Tsuji K, Bandyopadhyay A, Harfe BD, Cox K, Kakar S, Gerstenfeld L, Einhorn T, Tabin CJ, Rosen V (2006) BMP2 activity, although dispensable for bone formation, is required for the initiation of fracture healing. Nat Genet 38:1424–1429
Tsuji K, Cox K, Bandyopadhyay A, Harfe BD, Tabin CJ, Rosen V (2008) BMP4 is dispensable for skeletogenesis and fracture-healing in the limb. J Bone Joint Surg Am 90 (Suppl 1):14–18
Tsuji K, Cox K, Gamer L, Graf D, Economides A, Rosen V (2010) Conditional deletion of BMP7 from the limb skeleton does not affect bone formation or fracture repair. J Orthop Res 28:384–389
Tsujimura T, Kinoshita M, Abe M (2006) Response of rabbit skeletal muscle to tibial lengthening. J Orthop Sci 11:185–190
Ulici V, Hoenselaar KD, Agoston H, McErlain DD, Umoh J, Chakrabarti S, Holdsworth DW, Beier F (2009) The role of Akt1 in terminal stages of endochondral bone formation: angiogenesis and ossification. Bone 45:1133–1145
Ulsamer A, Ortuno MJ, Ruiz S, Susperregui AR, Osses N, Rosa JL, Ventura F (2008) BMP-2 induces Osterix expression through up-regulation of Dlx5 and its phosphorylation by p38. J Biol Chem 283:3816–3826
Urness LD, Sorensen LK, Li DY (2000) Arteriovenous malformations in mice lacking activin receptor-like kinase-1. Nat Genet 26:328–331
Valdimarsdottir G, Goumans MJ, Rosendahl A, Brugman M, Itoh S, Lebrin F, Sideras P, Dijke P ten (2002) Stimulation of Id1 expression by bone morphogenetic protein is sufficient and necessary for bone morphogenetic protein-induced activation of endothelial cells. Circulation 106:2263–2270
Verrier S, Meury TR, Kupcsik L, Heini P, Stoll T, Alini M (2010) Platelet-released supernatant induces osteoblastic differentiation of human mesenchymal stem cells: potential role of BMP-2. Eur Cell Mater 20:403–414
Vinals F, Lopez-Rovira T, Rosa JL, Ventura F (2002) Inhibition of PI3K/p70 S6K and p38 MAPK cascades increases osteoblastic differentiation induced by BMP-2. FEBS Lett 510:99–104
Violini S, Ramelli P, Pisani LF, Gorni C, Mariani P (2009) Horse bone marrow mesenchymal stem cells express embryo stem cell markers and show the ability for tenogenic differentiation by in vitro exposure to BMP-12. BMC Cell Biol 10:29
Vogt J, Traynor R, Sapkota GP (2011) The specificities of small molecule inhibitors of the TGFβ and BMP pathways. Cell Signal 23:1831–1842
Wagers AJ, Conboy IM (2005) Cellular and molecular signatures of muscle regeneration: current concepts and controversies in adult myogenesis. Cell 122:659–667
Waggett AD, Benjamin M, Ralphs JR (2006) Connexin 32 and 43 gap junctions differentially modulate tenocyte response to cyclic mechanical load. Eur J Cell Biol 85:1145–1154
Wagner DO, Sieber C, Bhushan R, Borgermann JH, Graf D, Knaus P (2010) BMPs: from bone to body morphogenetic proteins. Sci Signal 3:mr1
Wakitani S, Mitsuoka T, Nakamura N, Toritsuka Y, Nakamura Y, Horibe S (2004) Autologous bone marrow stromal cell transplantation for repair of full-thickness articular cartilage defects in human patellae: two case reports. Cell Transplant 13:595–600
Wallace AL, Makki R, Weiss JB, Hughes SP (1995) Measurement of serum angiogenic factor in devascularized experimental tibial fractures. J Orthop Trauma 9:324–332
Wang EA, Rosen V, D’Alessandro JS, Bauduy M, Cordes P, Harada T, Israel DI, Hewick RM, Kerns KM, LaPan P (1990) Recombinant human bone morphogenetic protein induces bone formation. Proc Natl Acad Sci USA 87:2220–2224
Wang XT, Liu PY, Tang JB (2004) Tendon healing in vitro: genetic modification of tenocytes with exogenous PDGF gene and promotion of collagen gene expression. J Hand Surg Am 29:884–890
Wang QW, Chen ZL, Piao YJ (2005) Mesenchymal stem cells differentiate into tenocytes by bone morphogenetic protein (BMP) 12 gene transfer. J Biosci Bioeng 100:418–422
Wang X, Goh CH, Li B (2007) p38 mitogen-activated protein kinase regulates osteoblast differentiation through osterix. Endocrinology 148:1629–1637
Wang CJ, Weng LH, Hsu SL, Sun YC, Yang YJ, Chan YS, Yang YL (2010a) pCMV-BMP-2-transfected cell-mediated gene therapy in anterior cruciate ligament reconstruction in rabbits. Arthroscopy 26:968–976
Wang H, Noulet F, Edom-Vovard F, Tozer S, Le GF, Duprez D (2010b) Bmp signaling at the tips of skeletal muscles regulates the number of fetal muscle progenitors and satellite cells during development. Dev Cell 18:643–654
Wang K, Feng H, Ren W, Sun X, Luo J, Tang M, Zhou L, Weng Y, He TC, Zhang Y (2011) BMP9 inhibits the proliferation and invasiveness of breast cancer cells MDA-MB-231. J Cancer Res Clin Oncol 137:1687–1696
Wen Z, Han L, Bamburg JR, Shim S, Ming GL, Zheng JQ (2007) BMP gradients steer nerve growth cones by a balancing act of LIM kinase and Slingshot phosphatase on ADF/cofilin. J Cell Biol 178:107–119
White AP, Vaccaro AR, Hall JA, Whang PG, Friel BC, McKee MD (2007) Clinical applications of BMP-7/OP-1 in fractures, nonunions and spinal fusion. Int Orthop 31:735–741
Wiley DM, Kim JD, Hao J, Hong CC, Bautch VL, Jin SW (2011) Distinct signalling pathways regulate sprouting angiogenesis from the dorsal aorta and the axial vein. Nat Cell Biol 13:686–692
Wolfman NM, Hattersley G, Cox K, Celeste AJ, Nelson R, Yamaji N, Dube JL, Blasio-Smith E, Nove J, Song JJ, Wozney JM, Rosen V (1997) Ectopic induction of tendon and ligament in rats by growth and differentiation factors 5, 6, and 7, members of the TGF-beta gene family. J Clin Invest 100:321–330
Wong YP, Fu SC, Cheuk YC, Lee KM, Wong MW, Chan KM (2005) Bone morphogenetic protein 13 stimulates cell proliferation and production of collagen in human patellar tendon fibroblasts. Acta Orthop 76:421–427
Xiao Q, Du Y, Wu W, Yip HK (2010) Bone morphogenetic proteins mediate cellular response and, together with Noggin, regulate astrocyte differentiation after spinal cord injury. Exp Neurol 221:353–366
Xiao C, Zhou H, Liu G, Zhang P, Fu Y, Gu P, Hou H, Tang T, Fan X (2011) Bone marrow stromal cells with a combined expression of BMP-2 and VEGF-165 enhanced bone regeneration. Biomed Mater 6:015013
Yamada M, Akeda K, Asanuma K, Thonar EJ, An HS, Uchida A, Masuda K (2008) Effect of osteogenic protein-1 on the matrix metabolism of bovine tendon cells. J Orthop Res 26:42–48
Yamaguchi K, Nagai S, Ninomiya-Tsuji J, Nishita M, Tamai K, Irie K, Ueno N, Nishida E, Shibuya H, Matsumoto K (1999) XIAP, a cellular member of the inhibitor of apoptosis protein family, links the receptors to TAB1-TAK1 in the BMP signaling pathway. EMBO J 18:179–187
Yee Lui PP, Wong YM, Rui YF, Lee YW, Chan LS, Chan KM (2011) Expression of chondro-osteogenic BMPs in ossified failed tendon healing model of tendinopathy. J Orthop Res 29:816–821
Yu Y, Bliss JP, Bruce WJ, Walsh WR (2007) Bone morphogenetic proteins and Smad expression in ovine tendon-bone healing. Arthroscopy 23:205–210
Yu YY, Lieu S, Lu C, Colnot C (2010) Bone morphogenetic protein 2 stimulates endochondral ossification by regulating periosteal cell fate during bone repair. Bone 47:65–73
Zaidi N, Nixon AJ (2007) Stem cell therapy in bone repair and regeneration. Ann N Y Acad Sci 1117:62–72
Zammit PS (2008) All muscle satellite cells are equal, but are some more equal than others? J Cell Sci 121:2975–2982
Zamurovic N, Cappellen D, Rohner D, Susa M (2004) Coordinated activation of notch, Wnt, and transforming growth factor-beta signaling pathways in bone morphogenic protein 2-induced osteogenesis. Notch target gene Hey1 inhibits mineralization and Runx2 transcriptional activity. J Biol Chem 279:37704–37715
Zanotti S, Smerdel-Ramoya A, Stadmeyer L, Canalis E (2008) Activation of the ERK pathway in osteoblastic cells, role of gremlin and BMP-2. J Cell Biochem 104:1421–1426
Zelzer E, Olsen BR (2005) Multiple roles of vascular endothelial growth factor (VEGF) in skeletal development, growth, and repair. Curr Top Dev Biol 65:169–187
Zelzer E, Mamluk R, Ferrara N, Johnson RS, Schipani E, Olsen BR (2004) VEGFA is necessary for chondrocyte survival during bone development. Development 131:2161–2171
Zhao M, Harris SE, Horn D, Geng Z, Nishimura R, Mundy GR, Chen D (2002) Bone morphogenetic protein receptor signaling is necessary for normal murine postnatal bone formation. J Cell Biol 157:1049–1060
Zou H, Wieser R, Massague J, Niswander L (1997) Distinct roles of type I bone morphogenetic protein receptors in the formation and differentiation of cartilage. Genes Dev 11:2191–2203
Acknowledgements
We thank Gerburg K. Schwaerzer for critically reading the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Karen Ruschke and Christian Hiepen contributed equally to the work.
The authors are supported by the German Research Foundation (BSRT fellowship to C.H. and J.B.) and in part by the Research Committee, Freie Universität Berlin (K.R.).
Rights and permissions
About this article
Cite this article
Ruschke, K., Hiepen, C., Becker, J. et al. BMPs are mediators in tissue crosstalk of the regenerating musculoskeletal system. Cell Tissue Res 347, 521–544 (2012). https://doi.org/10.1007/s00441-011-1283-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00441-011-1283-6